THE LIVING WORLD
Unit Three. The Continuity of Life
9.5. Evolutionary Consequences of Sex
As you can now appreciate, meiosis is a lot more complicated than mitosis. Why has evolution gone to so much trouble? While our knowledge of how meiosis and sex evolved is sketchy, it is abundantly clear that meiosis and sexual reproduction have an enormous impact on how species continue to evolve today because of their ability to rapidly generate new genetic combinations. Three mechanisms each make key contributions: independent assortment, crossing over, and random fertilization.
Independent Assortment
The reassortment of genetic material that takes place during meiosis is the principal factor that has made possible the evolution of eukaryotic organisms, in all their bewildering diversity, over the past 1.5 billion years. Sexual reproduction represents an enormous advance in the ability of organisms to generate genetic variability. To understand, recall that most organisms have more than one chromosome. For example, the organism represented in figure 9.10 has three pairs of chromosomes, each offspring receiving three homologues from each parent, purple from the father and green from the mother. The offspring in turn produces gametes, but the distribution of homologues into the gametes is completely random. A gamete could receive all homologues that are paternal in origin, as on the far left; or it could receive all maternal homologues, as on the far right, or any combination. Independent assortment alone leads to eight possible gamete combinations. In human beings, each gamete receives one homologue of each of the 23 chromosomes, but which homologue of a particular chromosome it receives is determined randomly. Each of the 23 pairs of chromosomes migrates independently, so there are 223 (more than 8 million) different possible kinds of gametes that can be produced.
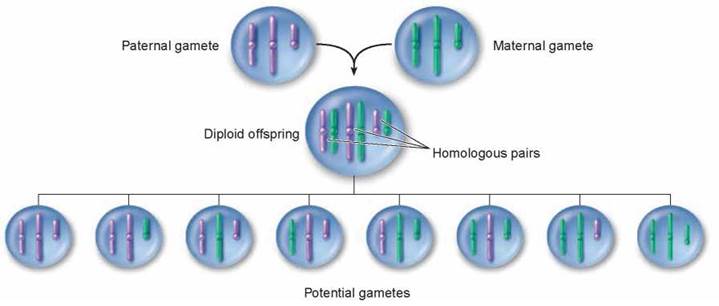
Figure 9.10. Independent assortment increases genetic variability.
Independent assortment contributes new gene combinations to the next generation because the orientation of chromosomes on the metaphase plate is random. In the cell shown here with three chromosome pairs, there are eight different gametes that can result, each with different combinations of parental chromosomes.
To make this point to his class, one professor offers an “A” course grade to any student who can write down all the possible combinations of heads and tails (an “either/or” choice, like that of a chromosome migrating to one pole or the other) with flipping a coin 23 times (like 23 chromosomes moving independently). No student has ever won an “A,” as there are over 8 million possibilities.
Crossing Over
The DNA exchange that occurs when the arms of nonsister chromatids cross over adds even more recombination. The number of possible genetic combinations that can occur among gametes is virtually unlimited.
Random Fertilization
Furthermore, because the zygote that forms a new individual is created by the fusion of two gametes, each produced independently, fertilization squares the number of possible outcomes (223 x 223= 70 trillion).
Importance of Generating Diversity
Paradoxically, the evolutionary process is both revolutionary and conservative. It is revolutionary in that the pace of evolutionary change is quickened by genetic recombination, much of which results from sexual reproduction. It is conservative in that change is not always favored by selection, which may instead preserve existing combinations of genes. These conservative pressures appear to be greatest in some asexually reproducing organisms that do not move around freely and that live in especially demanding habitats. In vertebrates, on the other hand, the evolutionary premium appears to have been on versatility, and sexual reproduction is the predominant mode of reproduction.
Key Learning Outcome 9.5. Sexual reproduction increases genetic variability through independent assortment in metaphase I of meiosis, crossing over in prophase I of meiosis, and random fertilization.
A Closser Look
Why Sex?
Not all reproduction is sexual. In asexual reproduction, an individual inherits all of its chromosomes from a single parent and is, therefore, genetically identical to its parent. Prokaryotic cells reproduce asexually, undergoing binary fission to produce two daughter cells containing the same genetic information.
Most protists reproduce asexually except under conditions of stress; then they switch to sexual reproduction. Among plants and fungi, asexual reproduction is common.
In animals, asexual reproduction often involves the budding off of a localized mass of cells, which grows by mitosis to form a new individual.
Even when meiosis and the production of gametes occur, there may still be reproduction without sex. The development of an adult from an unfertilized egg, called parthenogenesis, is a common form of reproduction in arthropods. Among bees, for example, fertilized eggs develop into diploid females, but unfertilized eggs develop into haploid males. Parthenogenesis even occurs among the vertebrates. Some lizards, fishes, and amphibians are capable of reproducing in this way; their unfertilized eggs undergo a mitotic nuclear division without cell cleavage to produce a diploid cell, which then develops into an adult. In some plants, such as hawkweeds, dandelions, and blackberries, a process similar to parthenogenesis called apomixis can occur.
If reproduction can occur without sex, why does sex occur at all? This question has generated considerable discussion, particularly among evolutionary biologists. Sex is of great evolutionary advantage for populations or species, which benefit from the variability generated in meiosis by random orientation of chromosomes and by crossing over. However, evolution occurs because of changes at the level of individual survival and reproduction, rather than at the population level, and no obvious advantage accrues to the progeny of an individual that engages in sexual reproduction. In fact, recombination is a destructive as well as a constructive process in evolution. The segregation of chromosomes during meiosis tends to disrupt advantageous combinations of genes more often than it creates new, better adapted combinations; as a result, some of the diverse progeny produced by sexual reproduction will not be as well adapted as their parents were. In fact, the more complex the adaptation of an individual organism, the less likely that recombination will improve it, and the more likely that recombination will disrupt it. It is, therefore, a puzzle to know what a well-adapted individual gains from participating in sexual reproduction, as all of its progeny could maintain its successful gene combinations if that individual simply reproduced asexually.
The DNA Repair Hypothesis. Several geneticists have suggested that sex occurs because only a diploid cell can effectively repair certain kinds of chromosome damage, particularly double-strand breaks in DNA. Both radiation and chemical events within cells can induce such breaks. As organisms became larger and longer-lived, it must have become increasingly important for them to be able to repair such damage. Synapsis, which in early stages of meiosis precisely aligns pairs of homologous chromosomes, may well have evolved originally as a mechanism for repairing double-strand damage to DNA. The undamaged homologous chromosome could be used as a template to repair the damaged chromosome. A transient diploid phase would have provided an opportunity for such repair. In yeast, mutations that inactivate the repair system for double-strand breaks of the chromosomes also prevent crossing over, suggesting a common mechanism for both synapsis and repair processes.
Muller's Ratchet. The geneticist Herman Muller pointed out in 1965 that asexual populations incorporate a kind of mutational ratchet mechanism—once harmful mutations arise, asexual populations have no way of eliminating them, and they accumulate over time, like turning a ratchet. Sexual populations, on the other hand, can employ recombination to generate individuals carrying fewer mutations, which selection can then favor. Sex may just be a way to keep the mutational load down.
The Red Queen Hypothesis. One evolutionary advantage of sex may be that it allows populations to "store” forms of a trait that are currently bad but have promise for reuse at some time in the future. Because populations are constrained by a changing physical and biological environment, selection is constantly acting against such traits. But in sexual species, selection can never get rid of those variants sheltered by more dominant forms of the trait.
The evolution of most sexual species, most of the time, thus manages to keep pace with ever-changing physical and biological constraints. This "treadmill evolution” is sometimes called the "Red Queen hypothesis,” after the Queen of Hearts in Lewis Carroll's Through the Looking Glass, who tells Alice, "Now, here, you see, it takes all the running you can do, to keep in the same place.”

Inquiry & Analysis
Are New Microtubules Made When the Spindle Forms?
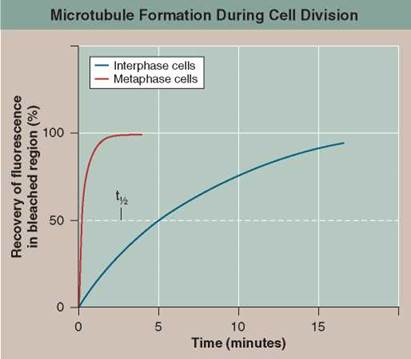
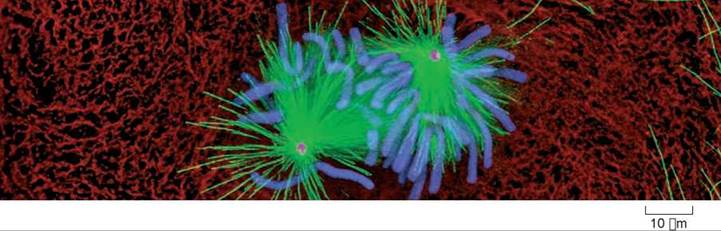
During interphase, before the beginning of meiosis, relatively few long microtubules extend from the centrosome (a zone around the centrioles of animal cells where microtubules are organized) to the cell periphery. Like most microtubules, these are refreshed at a low rate with resynthesis. Late in prophase, however, a dramatic change is seen—the centrosome divides into two, and a large increase is seen in the number of microtubules radiating from each of the two daughter centrosomes. The two clusters of new microtubules are easily seen as the green fibers connecting to the two sets of purple chromosomes in the micrograph of early prophase below (a micrograph is a photo taken through a microscope). This burst of microtubule assembly marks the beginning of the formation of the spindle characteristic of metaphase. When it first became known to cell biologists, they asked whether these were existing microtubules being repositioned in the spindle, or newly synthesized microtubules only produced just before metaphase begins.
The graph to the upper right displays the results of an experiment designed to answer this question. Mammalian cells in culture (cells in culture are growing in the laboratory on artificial medium) were injected with microtubule subunits (tubulin) to which a fluorescent dye had been attached (a fluorescent dye is one that glows when exposed to ultraviolet or short- wavelength visual light). After the fluorescent subunits had become incorporated into the cells's microtubules, all the fluorescence in a small region of a cell was bleached by an intense laser beam, destroying the microtubules there. Any subsequent rebuilding of microtubules in the bleached region would have to employ the fluorescent subunits present in the cell, causing recovery of fluorescence in the bleached region. The graph reports this recovery as a function of time, for interphase and metaphase cells. The dotted line represents the time for 50% recovery of fluorescence (t1/2) (that is, t1/2 is the time required for half of the microtubules in the region to be resynthesized).
1. Applying Concepts
a. Variable. In the graph, what is the dependent variable?
b. t1/2. Are new microtubules synthesized during interphase? What is the t1/2 of this replacement synthesis? Are new microtubules synthesized during metaphase? What is the t1/2 of this replacement synthesis?
2. Interpreting Data. Is there a difference in the rate at which microtubules are synthesized during interphase and metaphase? How big is the difference? What might account for it?
3. Making Inferences
a. What general statement can be made regarding the relative rates of microtubule production before and during meiosis?
b. Is there any difference in the final amount of microtubule synthesis that would occur if this experiment were to be continued for an additional 15 minutes?
4. Drawing Conclusions. When are the microtubules of the spindle assembled?
5. Further Analysis. The spindle breaks down after cell division is completed. Design an experiment to test whether the tubulin subunits of the spindle microtubules are recycled into other cell components, or destroyed, after meiosis.
Test Your Understanding
1. An egg and a sperm unite to form a new organism. To prevent the new organism from having twice as many chromosomes as its parents,
a. half of the chromosomes in the new organism quickly disassemble, leaving the correct number.
b. half of the chromosomes from the egg and half from the sperm are ejected from the new cell.
c. the large egg contains all the chromosomes, the tiny sperm only contributes some DNA.
d. the egg and sperm cells only have half the number of chromosomes found in the parents due to meiosis.
2. The diploid number of chromosomes in humans is 46. The haploid number is
a. 138.
b. 92.
c. 46.
d. 23.
3. In organisms that have sexual life cycles, there is a time when there are
a. 1n gametes (haploid), followed by 2n zygotes (diploid).
b. 2n gametes (haploid), followed by 1n zygotes (diploid).
c. 2n gametes (diploid), followed by 1n zygotes (haploid).
d. 1n gametes (diploid), followed by 2n zygotes (haploid).
4. Which of the following occurs in meiosis I?
a. All chromosomes duplicate.
b. Homologous chromosomes randomly orient themselves on the metaphase plate, called independent assortment.
c. The duplicated sister chromatids separate.
d. The original cell divides into four diploid cells.
5. Which of the following occurs in meiosis II?
a. All chromosomes duplicate.
b. Homologous chromosomes randomly separate, called independent assortment.
c. The duplicated sister chromatids separate.
d. Genetically identical daughter cells are produced.
6. During which stage of meiosis is crossing over initiated?
a. prophase I
b. anaphase I
c. metaphase II
d. interphase
7. Synapsis is the process whereby
a. homologous pairs of chromosomes separate and migrate toward a pole.
b. homologous chromosomes exchange chromosomal material.
c. homologous chromosomes become closely associated along their lengths.
d. the daughter cells contain half the number of chromosomes as the parent cell.
8. Crossing over is the process whereby
a. homologous chromosomes cross over to opposite sides of the cell.
b. homologous chromosomes exchange chromosomal material.
c. homologous chromosomes become closely associated along their lengths.
d. kinetochore fibers attach to both sides of a centromere.
9. Mitosis results in _____, while meiosis results in _____.
a. cells that are genetically identical to the parent cell/haploid cells
b. haploid cells/diploid cells
c. four daughter cells/two daughter cells
d. cells with half the number of chromosomes as the parent cell/cells that vary in chromosome number
10. A major consequence of sex and meiosis is that each species
a. remains pretty much the same because the chromosomes are carefully duplicated and passed to the next generation.
b. has a lot of genetic reassortment due to processes in meiosis II.
c. has a lot of genetic reassortment due to processes in meiosis I.
d. has a lot of genetic reassortment due to processes in telophase II.