Medical Microbiology
Section 1 The adversaries – microbes
1 Microbes as parasites
The varieties of microbes
Prokaryotes and eukaryotes
A number of important and distinctive biologic characteristics must be taken into account when considering any organism in relation to infectious disease. One of these is the way in which the organism is constructed, particularly the way in which genetic material and other cellular components are organized.
All organisms other than viruses and prions are made up of cells
Viruses are not cells – they do have genetic material (DNA or RNA) but lack cell membranes, cytoplasm and the machinery for synthesizing macromolecules, depending instead upon host cells for this process. Conventional viruses have their genetic material packed in capsules. The agents (prions) which cause diseases such as Creutzfeldt–Jakob disease (CJD), variant CJD and kuru in humans, scrapie and bovine spongiform encephalopathy (BSE) in animals, appear to lack nucleic acid and consist only of infectious proteinaceous particles.
All other organisms have a cellular organization, their bodies being made up of single cells (most ‘microbes’) or of many cells. Each cell has genetic material (DNA) and cytoplasm with synthetic machinery, and is bounded by a cell membrane.
Bacteria are prokaryotes, all other organisms are eukaryotes
There are many differences between the two major divisions: prokaryotes and eukaryotes, of cellular organisms (Fig. 1.1). These include the following.
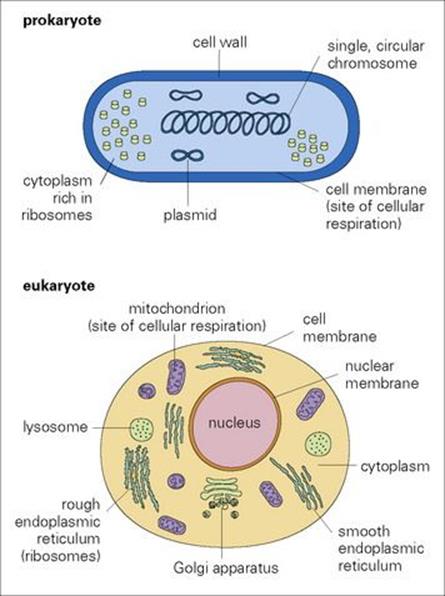
Figure 1.1 Prokaryote and eukaryote cells. The major features of cellular organization are shown diagrammatically.
In prokaryotes:
• A distinct nucleus is absent.
• DNA is in the form of a single circular chromosome. Additional ‘extrachromosomal’ DNA is carried in plasmids.
• Transcription and translation can be carried out simultaneously.
In eukaryotes:
• DNA is carried on several chromosomes within a nucleus.
• The nucleus is bounded by a nuclear membrane.
• Transcription requires formation of messenger RNA (mRNA) and movement of mRNA out of the nucleus into the cytoplasm
• Translation takes place on ribosomes.
• The cytoplasm is rich in membrane-bound organelles (mitochondria, endoplasmic reticulum, Golgi apparatus, lysosomes) which are absent in prokaryotes.
Gram-negative bacteria have an outer lipopolysaccharide-rich layer
Another important difference between prokaryotes and the majority of eukaryotes is that the cell membrane (plasma membrane) of prokaryotes is covered by a thick protective cell wall. In Gram-positive bacteria, this wall, made of peptidoglycan, forms the external surface of the cell, while in Gram-negative bacteria there is an additional outer layer rich in lipopolysaccharides. These layers play an important role in protecting the cell against the immune system and chemotherapeutic agents, and in stimulating certain pathologic responses. They also confer antigenicity.
Microparasites and macroparasites
Microparasites replicate within the host
There is an important distinction between microparasites and macroparasites that overrides their differences in size. Microparasites (viruses, bacteria, protozoa, fungi) replicate within the host and can, theoretically, multiply to produce a very large number of progeny, thereby causing an overwhelming infection. In contrast, macroparasites (worms, arthropods), even those that are microscopic, do not have this ability: one infectious stage matures into one reproducing stage, and, in most cases, the resulting progeny leave the host to continue the cycle. The level of infection is therefore determined by the numbers of organisms that enter the body. This distinction between microparasites and macroparasites has important clinical and epidemiologic implications.
The boundary between microparasites and macroparasites is not always clear. The progeny of some macroparasites do remain within the host, and infections can lead to the build-up of overwhelming numbers, particularly in immune-suppressed patients. The roundworms Trichinella,Strongyloides stercoralis and some filarial nematodes, and Sarcoptes scabiei (the itch mite), are examples of this type of parasite.
Organisms that are small enough can live inside cells
Absolute size has other biologically significant implications for the host–pathogen relationship, which cut across the divisions between micro- and macroparasites. Perhaps the most important of these is the relative size of a pathogen and its host’s cells. Organisms that are small enough can live inside cells and, by doing so, establish a biologic relationship with the host that is quite different from that of an extracellular organism – one that influences both disease and control.
Living inside or outside cells
The basis of all host–pathogen relationships is the exploitation by one organism (the pathogen) of the environment provided by another (the host). The nature and degree of exploitation varies from relationship to relationship, but the pathogen’s primary requirement is a supply of metabolic materials from the host, whether provided in the form of nutrients or (as in the case of viruses) in the form of nuclear synthetic machinery. The reliance of viruses upon host synthetic machinery requires an obligatory intracellular habit: viruses must live within host cells. Some other groups of pathogens (Chlamydia, Rickettsia) also live only within cells. In the remaining groups of pathogens, different species have adopted either the intracellular or the extracellular habit, or, in a few cases, both. Intracellular microparasites other than viruses take their metabolic requirements directly from the pool of nutrients available in the cell itself, whereas extracellular organisms take theirs from the nutrients present in tissue fluids, or, occasionally, by feeding directly on host cells (e.g. Entamoeba histolytica, the organism associated with amoebic dysentery). Macroparasites are almost always extracellular (though Trichinella is intracellular), and many feed by ingesting and digesting host cells; others can take up nutrients directly from tissue fluids or intestinal contents.
Pathogens within cells are protected from many of the host’s defence mechanisms
As will be discussed in greater detail in Chapter 13, the intracellular pathogens pose problems for the host that are quite different from those posed by extracellular organisms. Pathogens that live within cells are largely protected against many of the host’s defence mechanisms while they remain there, particularly against the action of specific antibodies. Control of these infections depends therefore on the activities of intracellular killing mechanisms, short-range mediators or cytotoxic agents, although the latter may destroy both the pathogen and the host cell, leading to tissue damage. This problem, of targeting activity against the pathogen when it lives within a vulnerable cell, also arises when using drugs or antibiotics, as it is difficult to achieve selective action against the pathogen while leaving the host cell intact. Even more problematic is the fact that many intracellular pathogens live inside the very cells responsible for the host’s immune and inflammatory mechanisms and therefore depress the host’s defensive abilities. For example, a variety of viral, bacterial and protozoal pathogens live inside macrophages, and several viruses (including HIV) are specific for lymphocytes.
Intracellular life has many advantages for the pathogen. It provides access to the host’s nutrient supply and its genetic machinery and allows escape from host surveillance and antimicrobial defences. However, no organism can be wholly intracellular at all times: if it is to replicate successfully, transmission must occur between the host’s cells, and this inevitably involves some exposure to the extracellular environment. As far as the host is concerned, this extracellular phase in the development of the pathogen provides an opportunity to control infection through defence mechanisms such as phagocytosis, antibody and complement. However, transmission between cells can involve destruction of the initially infected cell and so contribute to tissue damage and general host pathology.
Living outside cells provides opportunities for growth, reproduction and dissemination
Extracellular pathogens can grow and reproduce freely, and may move extensively within the tissues of the body. However, they also face constraints on their survival and development. The most important is continuous exposure to components of the host’s defence mechanisms, particularly antibody, complement and phagocytic cells.
The characteristics of extracellular organisms lead to pathologic consequences that are quite different from those associated with intracellular species. These are seen most dramatically with the macroparasites, whose sheer physical size, reproductive capacity and mobility can result in extensive destruction of host tissues. Many extracellular pathogens have the ability to spread rapidly through extracellular fluids or to move rapidly over surfaces, resulting in a widespread infection within a relatively short time. The rapid colonization of the entire mucosal surface of the small bowel by Vibrio cholerae is a good example. Successful host defence against extracellular parasites requires mechanisms that differ from those used in defence against intracellular parasites. The variety of locations and tissues occupied by extracellular parasites also poses problems for the host in ensuring effective deployment of defence mechanisms. Defence against intestinal parasites requires components of the innate and adaptive immune systems that are quite distinct from those effective against parasites in other sites, and those living in the lumen may be unaffected by responses operating in the mucosa. These problems in mounting effective defence are most acute where large macroparasites are concerned, because their size often renders them insusceptible to defence mechanisms that can be used against smaller organisms. For example, worms cannot be phagocytosed; they often have protective external layers, and can actively move away from areas where the host response is activated.
Systems of classification
Infectious diseases are caused by organisms belonging to a very wide range of different groups – prions, viruses, bacteria, fungi, protozoa, helminths (worms) and arthropods. Each has its own system of classification, making it possible to identify and categorize the organisms concerned. Correct identification is an essential requirement for accurate diagnosis and effective treatment. Identification is achieved by a variety of means, from simple observation to molecular analysis. Classification is being revolutionized by the application of genome sequencing. Many of the major pathogens in all categories have now been sequenced and this is allowing not only more precise identification but also a greater understanding of the interrelationships of members within each taxonomic group.
The approaches used vary between the major groups. For the protozoa, fungi, worms and arthropods, the basic unit of classification is the species, essentially defined as a group of organisms capable of reproducing sexually with one another. Species provide the basis for the binomial system of classification, used for eukaryote and some prokaryote organisms. Species are in turn grouped into a ‘genus’ (closely related but non-interbreeding species). Each organism is identified by two names, indicating the ‘genus’ and the ‘species’, respectively, for example, Homo sapiens andEscherichia coli. Related genera are grouped into progressively broader and more inclusive categories.
Classification of bacteria and viruses
The concept of ‘species’ is a basic difficulty in classifying prokaryotes and viruses, although the categories of genus and species are routinely used for bacteria. Classification of bacteria uses a mixture of easily determined microscopic, macroscopic and biochemical characteristics, based on size, shape, colour, staining properties, respiration and reproduction, and a more sophisticated analysis of immunologic and molecular criteria. The former characteristics can be used to divide the organisms into conventional taxonomic groupings, as shown for the Gram-positive bacteria inFigure 1.2 (see also Ch. 2).
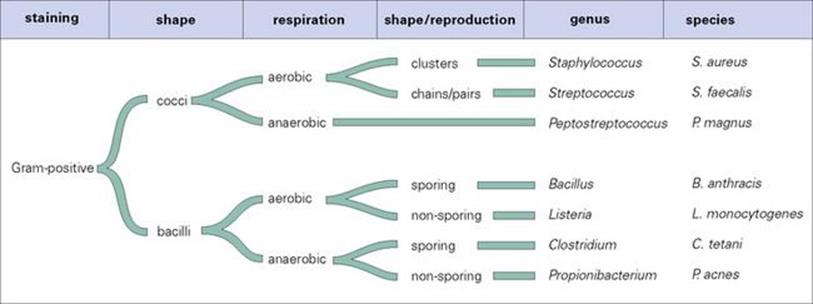
Figure 1.2 How the structural and biologic characteristics of bacteria can be used in classification, taking Gram-positive bacteria as an example.
Correct identification of bacteria below the species level is often vital to differentiate pathogenic and non-pathogenic forms
Correct treatment requires correct identification. For some bacteria, the important subspecies groups are identified on the basis of their immunologic properties. Cell wall, flagellar and capsule antigens are used in tests with specific antisera to define serogroups and serotypes (e.g. in salmonellae, streptococci, shigellae, E. coli). These tests are particularly useful for those organisms which grow poorly or not at all in vitro. Biochemical characteristics can be used to define other subspecies groupings (biotypes, strains, groups). For example, certain strains of Staphylococcus aureus release a β-haemolysin (causing red blood cells to lyse). Production of other toxins is also important in differentiating between groups, as in E. coli. Antibiotic susceptibility is also a useful technique for identification. Bacteria can also be classified below species level by their susceptibility to particular bacteriophage viruses. Phage typing is used, e.g. in differentiating between isolates of Vibrio cholerae and Salmonella enterica serovars.
Direct genetic approaches are also used in identification and classification such as the use of the polymerase chain reaction (PCR) and probes to detect organism-specific sentinel DNA sequences.
Classification of viruses departs even further from the binomial system
For viruses, families and, sometimes, genera are used, but there is much debate about the validity of the species concept for these organisms. Virus names draw on a wide variety of characteristics, e.g. size, structure, pathology, tissue location or distribution. Groupings are based on characteristics such as the type of nucleic acid present (DNA or RNA), the mode of replication, the symmetry of the virus particle (icosahedral, helical or complex) and the presence or absence of an external envelope, as shown for the DNA viruses in Figure 1.3 (see also Ch. 3). The equivalents of subspecies categories are also used, and indeed are more easily determined than species could be, given the peculiar biologic characteristics of viruses. These categories include serotypes, strains, variants and isolates and are determined primarily by serologic reactivity of virus material. The influenza virus, for example, can be considered as the equivalent of a genus containing three types (A, B, C). Identification can be carried out using the stable nucleoprotein antigen, which differs between the three types. The neuraminidase and haemagglutinin antigens are not stable and show variation within types. Characterization of these antigens in an isolate enables the particular variant to be identified, haemagglutinin (H) and neuraminidase (N) variants being designated by numbers, e.g. H5N1, the variant associated with fatal avian influenza (see Ch. 19). A further example is seen in adenoviruses, for which the various antigens associated with a component of the capsid can be used to define groups, types and finer subdivisions. The rapid rate of mutation shown by some viruses (e.g. HIV) creates particular problems for classification. The population present in a virus-infected individual may be genetically quite diverse and may best be described as a quasispecies – representing the average of the broad spectrum of variants present.
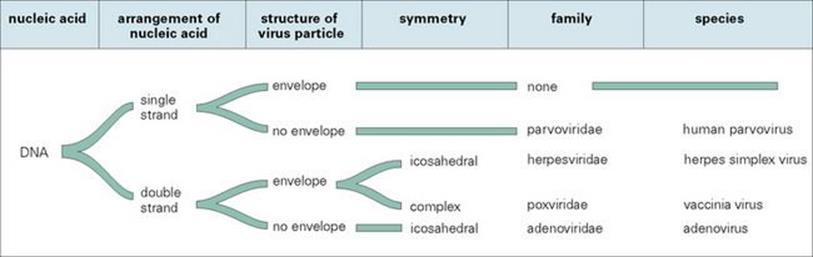
Figure 1.3 How the characteristics of viruses can be used in classification, taking DNA viruses as an example.
Classification assists diagnosis and the understanding of pathogenicity
Prompt identification of organisms is necessary clinically so that diagnoses can be made and appropriate treatments advised. To understand host–parasite interactions, however, not only should the identity of an organism be known, but as much as possible of its general biology; useful predictions can then be made about the consequences of infection. For these reasons, in subsequent chapters, we have included outline classifications of the important pathogens, accompanied by brief accounts of their structure (gross and microscopic), modes of life, molecular biology, biochemistry, replication and reproduction.
![]()
 Key Facts
Key Facts
• Organisms that cause infectious diseases can be grouped into seven major categories: prions, viruses, bacteria, fungi, protozoa, helminths and arthropods.
• Identification and classification of these organisms is an important part of microbiology and essential for correct diagnosis, treatment and control.
• Each group has distinctive characteristics (structural and molecular make-up, biochemical and metabolic strategies, reproductive processes) which determine how the organisms interact with their hosts and how they cause disease.
• Many pathogens live within cells, where they are protected from many components of the host’s protective responses.
![]()