Medical Microbiology
Section 5 Diagnosis and control
34 Protecting the host
vaccination
Introduction
Vaccines are one of the most effective public health tools. This chapter will review how vaccines work, and the vaccines in current use. However, although vaccination is very cost-effective as a public health measure, saving many lives (immunization saves an estimated 2.5 million deaths each year from diphtheria, tetanus, pertussis and measles), in 2008 an estimated 1.8 million people died from a vaccine-preventable disease, as a result of poor vaccine uptake. Many others die from infectious diseases such as HIV for which there is as yet no effective vaccine, so new vaccines are also needed (Table 34.1).
Table 34.1 Infectious agents that are major killers
|
Organism |
Disease |
Estimated annual deaths (millions) |
|
HIV |
AIDS |
1.8 |
|
Mycobacterium tuberculosis |
Tuberculosis |
1.7 |
|
Plasmodium spp. |
Malaria |
0.8 |
|
Total |
4.3 |
We currently lack effective vaccines against these organisms, although bacille Calmette–Guérin (BCG) vaccination can provide protection against disseminated forms of childhood tuberculosis, and pulmonary tuberculosis in some parts of the world. Most of the deaths from HIV are in Africa, and most of the deaths from malaria are in African children. Sources: Figures for 2009 from WHO.
Vaccination exploits the ability of the immune system to develop immunological memory, so that it can rapidly mobilize its forces to fight infection when required. Vaccines can be of different types, including live attenuated organisms, killed organisms, or subunit vaccines. Depending on the vaccine type, more than one dose may be needed to achieve or maintain optimal protection. Adjuvants are often required to increase immunity. A lot has been learnt about how to make a good vaccine, but for some vaccines we lack measurable correlates of protection. It is therefore no surprise that the development of new and more effective vaccines is a major area of research. Successful vaccination also requires an understanding of the epidemiology of disease transmission, for example, to estimate what proportion of a given population needs to be vaccinated to produce herd immunity, as discussed in Chapter 31.
Vaccination – a four hundred year history
‘Never in the history of human progress’, wrote the pathologist Geoffrey Edsall, ‘has a better and cheaper method of preventing illness been developed than immunization at its best’. The greatest success story in medicine, the elimination of smallpox, began before either immunology or microbiology were recognized as disciplines – indeed before the existence of microbes or the immune system was even suspected. As a result of the pioneering work of Jenner with vaccinia (Box 34.1), all forms of specific, actively induced immunity are now referred to as ‘vaccination’.
![]()
Box 34.1  Lessons in Microbiology
Lessons in Microbiology
Edward Jenner (1749–1823)
The English physician Edward Jenner (Fig. 34.1) is regarded as the founder of modern vaccination, but he was by no means the first to try the technique. The ancient practice of ‘variolation’ dates back to tenth-century China, and arrived in Europe in the early eighteenth century via Turkey. The technique involved the inoculation of children with dried material from healed scabs of mild smallpox cases, and was a striking foretaste of the principles of modern attenuated viral vaccines. This practice was, however, both inconsistent and dangerous, and Jenner’s innovation was to show that a much safer and more reliable protection could be obtained by deliberate inoculation with cowpox (vaccinia) virus. Milkmaids exposed to cowpox were traditionally known to be resistant to smallpox and so retained their smooth complexions. In 1796, Jenner tested his theory by inoculating 8-year-old James Phipps with liquid from a cowpox pustule on the hand of Sarah Nelmes. Subsequent inoculation of the boy with smallpox produced no disease. Although greeted with scepticism at first, Jenner’s ideas soon became accepted, and he went on to inoculate thousands of patients in a shed in the garden of his house at Berkeley, Gloucestershire. He ultimately achieved world fame, though his fellowship of the Royal Society was conferred for a quite different piece of work on the nesting habits of the cuckoo!
![]()

Figure 34.1 Edward Jenner (1749–1823).
The principle of vaccination is simple: to prime the adaptive immune system to the antigens of a particular microbe so that on first contact with the live organism a rapid and effective secondary immune response will be induced by memory T and B cells. Vaccination therefore depends upon the ability of lymphocytes, both B and T cells, to respond to specific antigens and develop into memory T and B cells, and represents a form of actively enhanced adaptive immunity. The passive administration of preformed elements, such as antibody, is considered in Chapter 35.
Aims of vaccination
The aims of vaccination can vary from preventing symptoms to eradication of disease
The most ambitious aim of vaccination is eradication of the disease. This has been achieved for smallpox, the eradication of polio is being attempted, and there has clearly been a dramatic downward trend in the incidence of most of the diseases against which vaccines are currently in use (Fig. 34.2). However, as long as any focus of infection remains in the community, the main effect of vaccination will be protection of the vaccinated individual against infection.
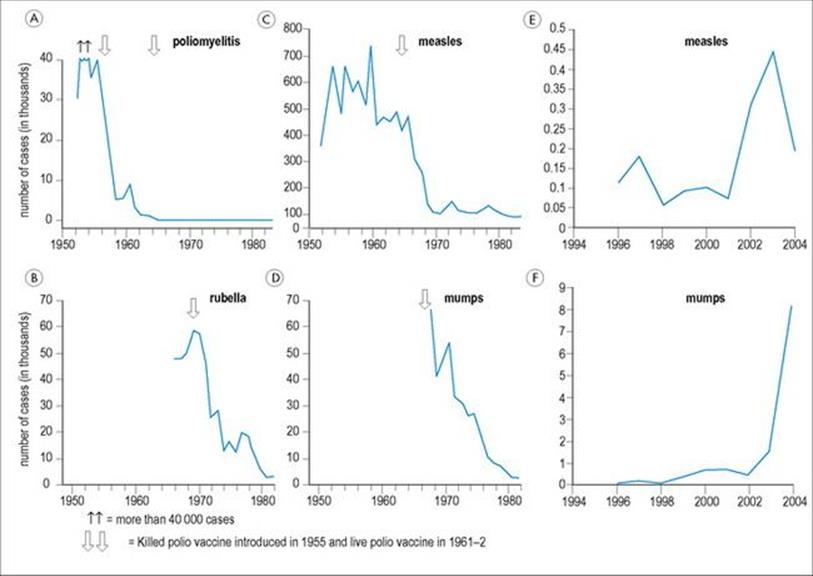
Figure 34.2 The effect of vaccination on the incidence of various viral diseases in the USA and the UK. Most infections have shown a dramatic downward trend after the introduction of a vaccine (arrows), but the right-hand panels show the resurgence in disease when vaccine uptake is reduced following vaccine ‘scares’.
(Data from Mims and White and the Health Protection Agency, UK.)
In certain cases, the aim of vaccination may be more limited: namely, to protect the individual against symptoms or pathology. For example, diphtheria and tetanus vaccines only induce immunity against the toxins produced by the bacteria, as it is the effect of these toxins rather than the simple presence of the microbe itself that is harmful.
The importance of herd immunity
Successful vaccination programmes rely not only on the development and use of vaccines themselves, but also on an understanding of the epidemiologic aspects of disease transmission. If enough individuals in a population are immunized, this will reduce or stop transmission of the infection. This is called herd immunity. By having your own child immunized, you therefore help protect the whole community – but conversely, when too many parents decide that their child will not be immunized, because they think the risk of their child getting the disease is low, this may contribute to the disease becoming more common (Fig. 34.2)
It is therefore important to know how many individuals in a population must be immunized to produce herd immunity, and also whether immunity should be boosted by re-vaccination.
Vaccines can be of different types
Vaccines can be based on whole organisms, either live or inactivated, or components of the infectious agent (Table 34.2).
Table 34.2 Types of vaccine
|
Types of vaccine |
Examples |
|
Live attenuated |
|
|
Viral |
Measles, mumps, rubella, vaccinia, varicella, yellow fever, zoster, oral polio, intranasal influenza, rotavirus |
|
Bacterial |
BCG, oral typhoid |
|
Inactivated |
|
|
Whole virus |
Polio, influenza, hepatitis A, rabies |
|
Whole bacteria |
Pertussis, cholera, typhoid |
|
Fractions |
|
|
Toxoids |
Diphtheria, tetanus |
|
Protein subunits |
Hepatitis B, influenza, acellular pertussis, human papillomavirus |
|
Polysaccharides |
Pneumococcal, meningococcal, Salmonella typhi (Vi), Haemophilus influenzae type b |
|
Conjugates |
Haemophilus influenzae type b, pneumococcal, meningococcal |
Note that not all types of vaccine are available in all countries. Vaccines are also available for bioterrorism agents such as anthrax and plague, and for vaccinia.
Live vaccines are designed to induce immunity in a similar way to the actual infection. Most live vaccines use attenuated organisms that were attenuated using culture in eggs, animals or in tissue culture (Fig. 34.3); these attenuated organisms replicate to a limited extent in the vaccinated individual but do not cause disease in healthy people. However, the immunosuppression associated with HIV infection can produce problems with live vaccines. For example, infants with HIV infection given BCG vaccination can develop disseminated BCGosis. HIV-infected individuals with severe immunosuppression should not be given live vaccines such as measles or varicella, but they can be given inactivated vaccines.
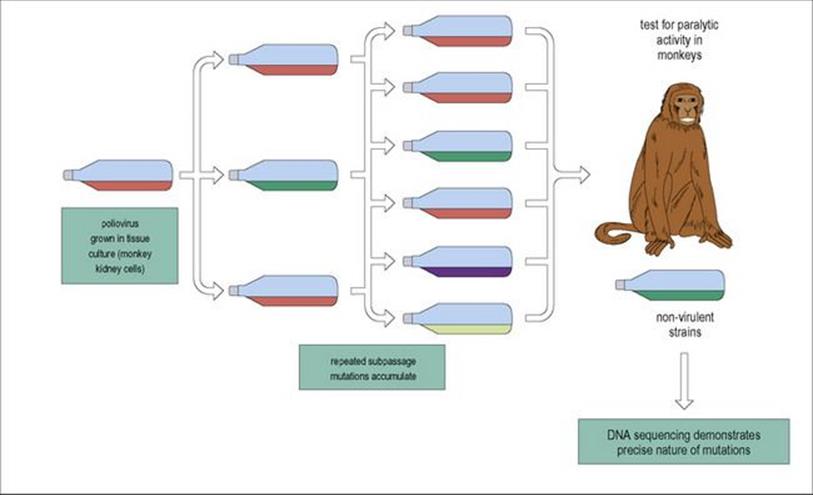
Figure 34.3 Live attenuated vaccines (e.g. polio) were originally produced by allowing viruses to grow in unusual conditions, and selecting the randomly occurring mutants that had lost virulence.
Inactivated vaccines are safe to use in the immunocompromised, although they may not be as immunogenic, and a good adjuvant may be needed. Inactivation can use heat or fixation, for example, with formalin. Types of fixatives and preservatives in use in vaccines are given in Table 34.3. Another difference between live and attenuated vaccines is that inactivated vaccines are not affected by circulating antibody, but live attenuated vaccines may be.
Table 34.3 Fixatives and preservatives used in current vaccines
|
Fixatives |
|
|
Formalin |
TdaP, HepA, Hib, influenza, polio |
|
Glutaraldehyde |
DtaP, DtaP-HepB-IPV |
|
Preservatives |
|
|
EDTA |
Rabies, varicella |
|
Phenol |
Pneumococcal, inactivated typhoid |
|
2-phenoxyethanol |
DtaP, HepA, HepA-HepB, inactivated poliovirus |
|
β-propiolactone |
Rabies |
|
Sodium deoxycholate |
Influenza |
|
Thiomersal |
DtaP, DtaP-Hib, HepB, influenza, meningococcal vaccine |
TdaP, combined tetanus, diphtheria and pertussis; DtaP, combined diphtheria, tetanus and pertussis; HepA, hepatitis A; HepB, hepatitis B; Hib, Hameophilus influenzae type b; IPV, inactivated polio vaccine.
Individual antigens or toxins can also be used as a vaccine, with adjuvant. Purified proteins are used in the acellular pertussis vaccine and recombinant proteins are used in the vaccines for hepatitis B. A number of protein antigens can also be joined together as a fusion protein and given with adjuvant. Polysaccharides form the basis of the pneumococcal vaccine, but as polysaccharide vaccines are not immunogenic in children under 2 years of age, conjugate vaccines that use a polysaccharide linked to a protein have been developed for pneumococcal and meningococcal disease, and for Haemophilus influenzae type b (Hib). For some bacteria, it is the toxin that is pathogenic – so this can be inactivated to make a toxoid, as in the tetanus toxoid vaccine. If individual components of an organism form the basis of a vaccine, an adjuvant will be needed to boost immune responses. Multiple doses of protein or polysaccharide are usually needed, as these vaccines are less immunogenic than vaccines with whole organisms.
One or more vaccine antigens can also be delivered by a viral vector, such as modified vaccinia virus Ankara (MVA) that was safely used in humans at the end of the smallpox eradication campaign.
Some vaccines are designed to boost immunity using only selected antigens or by using a different delivery route. For example, some new TB vaccines in development would boost the immunity induced by BCG by giving key antigens delivered by a viral vector (Fig. 34.4), or a fusion protein with adjuvant.
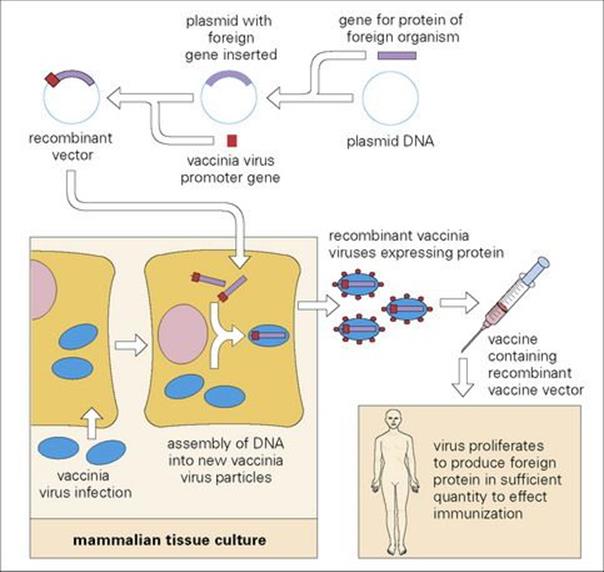
Figure 34.4 It is now possible to insert genes coding for antigens of one or more microorganisms into a large virus such as modified vaccinia virus Ankara (MVA), so that they replicate and are released into the host. A new MVA vaccine is currently being developed for TB that expresses a secretory TB protein.
Recipients of haemopoietic stem cell transplants may need to be revaccinated after the infusion of haemopoietic stem cells, as otherwise antibody titres to vaccine-preventable diseases decline.
Adjuvants
Adjuvants increase the immunity induced by a vaccine in a number of ways. Adjuvants can improve the immune response to the vaccine antigens through inducing activation of Toll-like receptors (TLR) on dendritic cells to improve antigen presentation, or by forming an antigen depot which allows antigen to persist and to leak out slowly over time. The earliest adjuvants consisted of water in oil emulsions, and Freund’s complete adjuvant which included dead mycobacteria in a water in oil emulsion is very effective, although not suitable for use in humans. Other adjuvants increase antigen presentation, or enhance particular types of immunity, such as antibodies or Th1 immunity. The dramatic effect of adding an adjuvant to a vaccine is shown in Figure 34.5. Aluminium salts are powerful adjuvants and used in many vaccines (Box 34.2). Experimentally, cytokines such as IL-1, IL-2, IFNγ, IL-12 and IL-18, as well as some chemokines, have been tested as adjuvants. Compounds such as liposomes, lipid containing vesicles, have also been used, but so far only monophosphoryl lipid A has been used in a licensed vaccine (HPV, Cervarix).
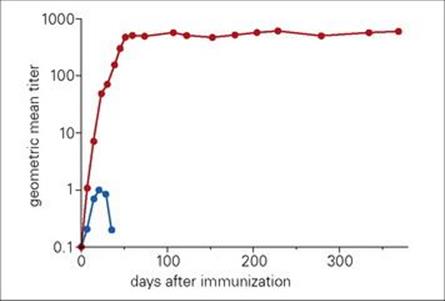
Figure 34.5 Effects of adjuvants on antibody responses of mice to egg albumin. Mice were injected subcutaneously with egg albumin in saline or in Freund’s incomplete adjuvant. Antibody titres at intervals over time are shown. The blue symbols represent antigen in saline, and the red symbols antigen in adjuvant.
(Redrawn from: Hunter, R. (2002) Vaccine 20:S7–S12.)
![]()
Box 34.2 Adjuvants in Currently Used Vaccines
|
Aluminium salts* |
DTaP, Hib, HepA, HepB, HPV, MMR, rabies, rotavirus, rubella, varicella |
|
Monophosphoryl lipid A (MPL) |
HPV (Cervarix) |
* aluminium hydroxide, aluminium hydroxyphosphate sulphate, aluminium phosphate, aluminium potassium sulphate
![]()
Vaccine safety
As vaccines are given to healthy individuals, it is important that they are safe. In 1926, live M. tuberculosis was inadvertently given to healthy children instead of BCG, leading to the Lubeck disaster, and in 1942, US military personnel were vaccinated with yellow fever virus contaminated with hepatitis B virus (see Ch. 15). Safety testing is now rigorous, requiring extensive quality controls and animal testing, prior to trials or use in humans. Some of the more important issues are summarised in Box 34.3. It is particularly critical that vaccines derived from live organisms are inactivated to ensure they are safe and that vaccines are preserved appropriately to ensure that vaccine immunogenicity is retained. Examples of fixatives and preservatives used in current vaccines are given in Table 34. 3.
![]()
Box 34.3 Problems with Vaccine Safety
Both living and non-living vaccines require rigorous quality and safety control. Some of the more common problems are listed below:
Live attenuated vaccines
• Insufficient attenuation
• Reversion to wild type
• Administration to immunodeficient patient
• Persistent infection
• Contamination by other viruses
• Fetal damage
Non-living vaccines
• Contamination by toxins or chemicals
• Allergic reactions
• Autoimmunity
Genetically engineered vaccines
• Possible inclusion of oncogenes
![]()
Vaccines in current use
Diphtheria, tetanus and pertussis
The diphtheria vaccine consists of the inactivated toxoid. Toxigenic Corynebacterium diphtheriae is grown in liquid culture and the filtrate inactivated with formaldehyde to produce the toxoid. This is a highly effective vaccine, giving > 90% protection. Three or four doses are required to give good protection, with a booster every 10 years. It is now given in different formulations with other vaccines.
The inactivated toxin from Clostridium tetani, inactivated using formaldehyde, is used to vaccinate against tetanus. Tetanus toxoid was first produced in 1924. Again, this is a very effective vaccine, but boosters are required every 10 years. In some developing countries neonatal tetanus is still a problem; if the mother has been immunized against tetanus this will protect the newborn baby but, in 2009, the World Health Organization estimated that only 83% of newborn babies were protected against neonatal tetanus.
The first vaccine developed against pertussis was a whole cell vaccine, which was available from the mid 1940s and introduced in the UK in 1957 (Fig. 34.6). However, although four doses of vaccine induced 70–90% protection against serious whooping cough, concerns over the safety of the vaccine in the UK in the 1970s led to resurgence of disease. Whether the vaccine did induce brain damage is unclear, but concerns did lead to the development of a safer acellular pertussis vaccine.
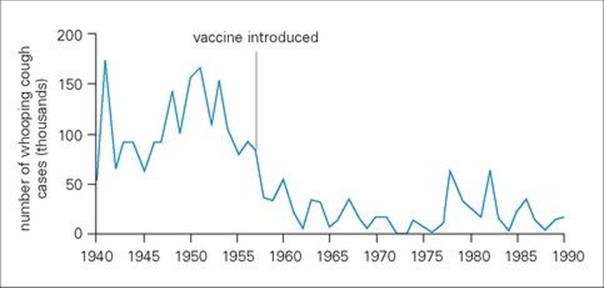
Figure 34.6 The number of cases of whooping cough notified fell steadily after the introduction of mass immunization in the UK in 1958, although epidemics continued to occur at approximately four-year intervals. Following the scare about the possible adverse effects of pertussis vaccine, the number of cases rose, and there was a large epidemic in the winter of 1978–79.
Coverage of the combined DTP or diptheria tetanus acellular pertussis (DTaP) vaccines is now good, with an estimated 109 million children under the age of 1 year receiving a third dose of vaccine, equivalent to 82% global coverage.
Measles, mumps and rubella vaccines
Live attenuated measles vaccine was introduced in the USA in 1963, using the Edmonston B vaccine, which has since been replaced by the more attenuated Edmonston–Enders strain, which is grown in tissue culture. Children should be given two doses of vaccine, as the first dose fails to induce protective antibodies in about 15% of those vaccinated. Vaccination is safe and effective, either given on its own or as part of the MMR vaccine with mumps and rubella and mumps, or the MMRV vaccine containing measles, mumps, rubella and varicella. However, maternal antibodies inhibit the induction of immunity, so the first dose is generally given at 12–15 months of age, once maternal-derived antibodies have declined, and the second at 4–6 years of age. In lower-income countries where the risk of contracting measles is higher, the vaccine may be given at about 9 months, in an attempt to protect children whose levels of maternal antibodies are declining.
Vaccine-induced immunity to measles is long-lived and after two doses, probably life-long. Between 2000 and 2008, there was a 78% drop in measles deaths worldwide, as a result of measles vaccination. In this period, approximately 700 million children were vaccinated, with 83% of all children vaccinated before 1 year of age in 2008. Nevertheless, it was estimated that about 450 children were still dying every day as a result of measles – measles is the leading cause of death in young children from a vaccine-preventable disease – and as many as 10 million people still catch measles every year. In the UK there were 1144 cases of measles in 2009 – with some clusters of cases in schools or nurseries.
As shown in Figure 34.2, cases of measles increased in the UK after 2001, following reduced vaccine uptake. This resulted from the suggestion that the MMR (measles, mumps and rubella) vaccine caused autism, as there was an apparent rise in autism in both California and the UK that seemed to coincide with the introduction of the MMR vaccine. However, further studies have failed to show an increased risk of autism after MMR. It had also been suggested that the measles virus could be detected in the gut following vaccination and that this might lead to inflammatory bowel disease – but, again, this was not confirmed by independent testing. It is no wonder that parents get worried when bombarded with these scare stories – but they forget that measles infection can kill healthy children. In a measles outbreak in Ireland in 2000, nearly 1500 cases were notified and three children died.
Mumps vaccine
The current mumps vaccine is a live attenuated virus (Jereyl Lynn strain), which was licensed in 1967. Over 97% of those vaccinated make antibodies after a single dose of vaccine, and a study in the UK showed that 88% of those receiving two doses were protected. The importance of receiving two doses of MMR is illustrated by a mumps outbreak in Northern Ireland where 55.4% of the confirmed cases had received one dose of vaccine, compared with 0.9% in those who had received two doses. After two doses, protection is thought to last at least 25 years and may be life-long. This vaccine is much more effective than an earlier inactivated vaccine.
Rubella vaccine
The current rubella vaccine is a live attenuated virus, strain 27/3, licensed in 1979. The virus was attenuated by 25–30 cell culture passages in human diploid fibroblasts. Over 90% of those vaccinated have at least 15 years of protection from clinical rubella or viraemia. Although rubella itself is a relatively mild infection, it causes real problems if pregnant woman become infected in the first trimester of pregnancy, when congenital rubella syndrome can cause serious damage to the fetus. Thankfully, there has been a dramatic reduction in confirmed cases of congenital rubella syndrome due to vaccination: cases were reduced by 98% in the Americas between 1998 and 2009.
Although a combined MMRV vaccine with measles, mumps, rubella and varicella is available, the risk of post-vaccination seizures seems slightly higher if this vaccine is given as the first vaccine dose at 12–15 months of age compared to giving the MMR and varicella vaccines separately. The combined vaccine can be given as the second dose at 4–6 years of age.
Polio vaccine
The first polio vaccine was a killed vaccine (inactivated polio vaccine, IPV) developed by Salk; this was first licensed in 1955, and was very effective at reducing the risk of contracting polio. Later, the oral polio vaccine (OPV) was developed by Sabin, and licensed in the 1960s. Giving the vaccine on sugar lumps or directly into the mouth was much easier than giving it by injection and the live vaccine also gives better intestinal immunity. However, the live poliovirus used in OPV vaccine is not genetically stable and can cause vaccine-associated paralytic polio (VAPP) in approximately 1 per million doses administered (Table 34.4). It has long been recognized that OPV is transmissible from vaccinees to their close contacts, and it is now known that such transmission can persist in the community as circulating vaccine-derived polio viruses (cVDPV). Extensive use of trivalent OPV in routine services and national immunization day campaigns, organized through the Global Poliovirus Eradication Initiative, has succeeded in reducing the number of polio cases worldwide by over 99%, from an estimated 350 000 cases in 1988 to only 1783 reported cases in 2009 (Fig. 34.7). Of the three wild polio serotypes, type 2 transmission was interrupted in 1999, but types 1 and 3 have continued to circulate in Afghanistan, India, Nigeria and Pakistan, and have caused outbreaks in some neighbouring countries.
Table 34.4 Oral and inactivated polio vaccines compared
|
Inactivated (IPV) |
Attenuated (OPV) |
|
|
Introduced |
Salk 1954 |
Sabin 1957 |
|
Route |
Injection plus alum |
Oral |
|
Advantages |
Can be given with other childhood vaccines |
Boosts IgA immunity |
|
Disadvantages |
Risk if inadequately killed |
Reversion to virulence* |
* Although vaccine-associated paralytic polio only occurs in < 1/million vaccinated, vaccine-derived polio viruses can circulate within the community.
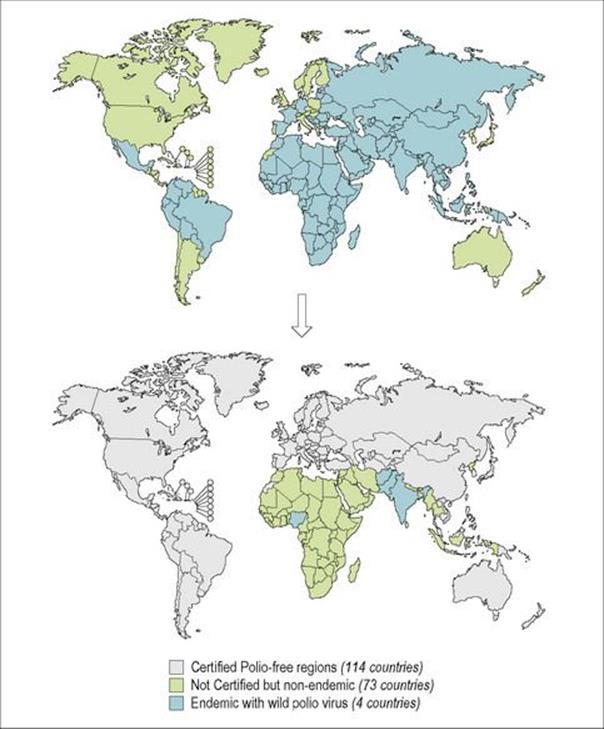
Figure 34.7 Progress towards polio eradication. The progress towards the eradication of polio is illustrated by the increase in certified polio-free countries from 1988 (top map) to 2009 (bottom map).
(Redrawn from www.who.int/immunization_monitoring/data/SlidesGlobalImmunization.pdf; data from WHO/Polio database, as at August 2010.)
Low effectiveness of trivalent vaccines in some populations (in particular India), and the elimination of type 2 virus, has led to the development of mono and bivalent vaccines with types 1 and 3 Sabin viruses. These are now widely employed in the Global Poliovirus Eradication Initiative. However to eradicate polio, the world may have to shift to IPV to avoid circulation of vaccine-derived polio viruses. Most wealthy countries have already shifted to IPV vaccines in recent years for this reason. The routine IPV vaccination schedule in the United States consists of four doses given at ages 2 months, 4 months, 6–18 months, and 4–6 years.
Pneumococcal vaccines
The challenge in making an effective vaccine against pneumococcal disease is that there are 90 serotypes of Streptococcus pneumoniae – but luckily a few serotypes cause most infections. The first vaccine was a pneumococcal polysaccharide vaccine with capsular polysaccharide from 14 serotypes. This was replaced in 1983 with a formulation containing 23 capsular polysaccharides from 23 serotypes. However, although this vaccine induced antibodies in > 80% of adults, it was not immunogenic in children aged less than 2 years. A conjugate vaccine is now available in which capsular polysaccharides from seven serotypes are conjugated to a non-toxic form of the diphtheria toxin. The conjugate vaccine is highly immunogenic in infants and young children, and new formulations containing more serotypes are being developed. One interesting question is whether the rates of carriage of the different serotypes may be affected by vaccination.
Meningococcal vaccines
As for pneumococcal vaccine, the first vaccine against meningococcal disease consisted of purified capsular polysaccharides for four of the five serotypes, A, C, Y and W-135. Again, similar to the pneumococcal polysaccharide vaccine, the meningococcal polysaccharide vaccine was not immunogenic in young children, as seen for other T-independent antigens. A conjugate vaccine containing capsular polysaccharides from the same four serotypes conjugated to diphtheria toxoid was licensed in the USA in 2005, and shown in trials to induce four times more antibody than the polysaccharide vaccine. The B strain is not covered in either of these vaccines, as the B group polysaccharide is poorly immunogenic and may have some cross-reactivity to the human nervous system. A vaccine containing just the C serotype is currently used in the UK. Vaccination against meningitis is compulsory for pilgrims visiting Mecca in Saudi Arabia for the Umrah and Haj pilgrimages, as there was a N. meningitis W-135 outbreak in pilgrims in 2000.
Haemophilus influenzae type b (Hib)
Haemophilus influenzae mainly affects children under 5 years of age. Although there are six capsular serotypes, one, type b composed of a phoshodiester-linked polymer of ribose and ribitol, causes 95% of disease, and so has been the basis of Hib vaccines. The introduction of Hib vaccines has dramatically reduced the incidence of bacterial meningitis from Hib (Fig. 34.8). The first polysaccharide vaccine introduced in the USA in 1985 was not immunogenic in children under 18 months of age, similar to other antigens inducing T cell-independent immune responses. Conjugating the polysaccharide to a T cell-dependent antigen overcomes this problem. Even so, three or four doses are needed to induce good immunity, as this is another example of how a subunit vaccine is less immunogenic than a live vaccine. Two conjugate vaccines are currently available, one conjugated to tetanus toxoid and one to meningococcal group B outer membrane protein. Another Hib capsular polysaccharide vaccine conjugated to inactivated tetanus toxoid is now available and can be used as the last booster dose.
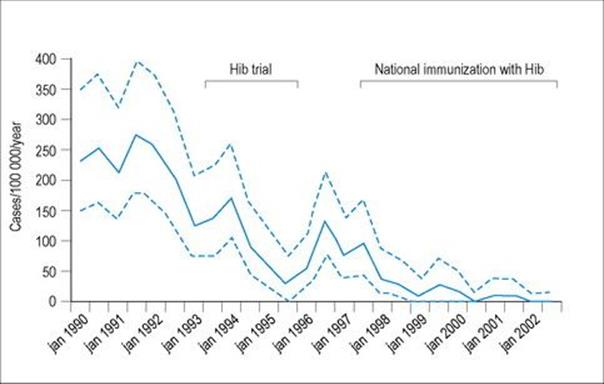
Figure 34.8 Vaccination with a Haemophilus influenzae type b (Hib) polysaccharide-tetanus toxoid conjugate vaccine has produced a dramatic decrease in the incidence of Hib meningitis in children > 1 year old in The Gambia. Dotted lines represent pointwise 90% likelihood-based confidence limits.
(Data from: Adegbola, R. et al. (2005) Lancet 366:144–150.)
Influenza
Flu generated a lot of alarm in 2009, when the first flu pandemic since 1968 was caused by a new influenza A (H1N1) virus. The threat from this new virus, and from avian influenza (H5N1), has highlighted the limited world capacity to produce new flu vaccines quickly in the quantities needed. Another result of the 2009 pandemic was a change to vaccination policies for flu in countries such as the USA. Two types of vaccine are currently available: a trivalent inactivated vaccine that can be given to anyone over the age of 6 months by intramuscular injection, and a live attenuated influenza vaccine, that can be given by intranasal spray to those aged 2–49 years of age who are healthy and not pregnant.
Flu is a tricky customer, as it changes its antigens due to both point mutations and to recombination events, resulting in antigenic drift (Fig. 34.9) and antigenic shift. The recommended composition of current flu vaccines can be found on the WHO website. The 2010–11 trivalent vaccines for the northern hemisphere contained A/California/7/2009 (H1N1)-like, A/Perth/16/2009 (H3N2)-like, and B/Brisbane/60/2008-like antigens. The influenza A (H1N1) vaccine virus was derived from a 2009 pandemic influenza A (H1N1) virus. Different formulations can be used for the southern hemisphere. Flu vaccination policy varies in different countries: for example, in 2010 in the USA, vaccination was offered to everyone aged over 6 months, whereas in the UK vaccination was restricted to those aged over 65 years or who fell into an at-risk group, such as those with asthma, but for the first time vaccination was offered to previously unvaccinated pregnant women. Children aged 6 months to 6 years (in the USA) or 12 years (in the UK) who are being vaccinated for the first time are now given two doses of vaccine. A new high-dose trivalent inactivated vaccine is also available for use in those over 65 years of age.
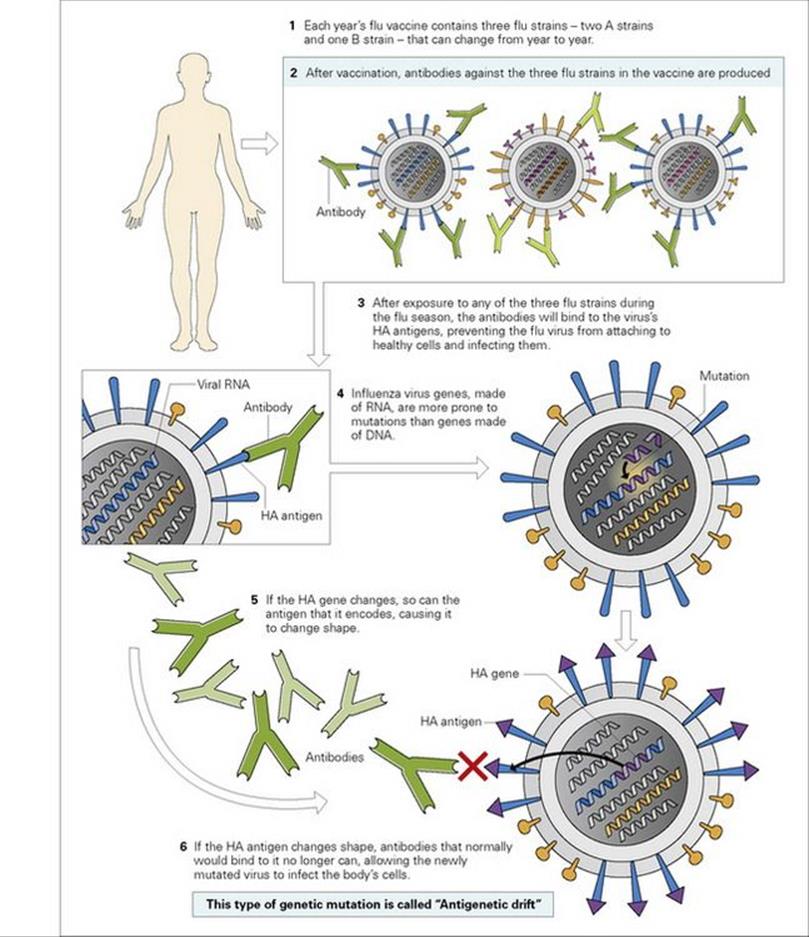
Figure 34.9 Influenza vaccines and antigenic drift. Seasonal influenza vaccines contain three flu strains, two A strains and one B strain. Antibodies to these strains induced by vaccination will protect against infection, but mutations in the influenza genes can cause antigenic drift leading to infection.
(Modified from National Institute of Allergy and Infectious Diseases. Flu (Influenza): Antigenic Drift. Bethesda, MD: U.S. Department of Health and Human Services; 2011.) HA haemagglutinin.
BCG and new vaccines for TB
The oldest vaccine still in use is the BCG vaccine, derived following extensive culture of M. bovis on potato bile medium by Calmette and Guérin, which resulted in its attenuation. The genetic deletions that this involved have since been mapped, and include the loss of the RD1 region that encodes the ESAT-6 and CFP-10 antigens that are used in the two currently available commercial diagnostic tests for M. tuberculosis infection, the QuantiFERON test and the TSPOT-TB ELISPOT assay. A number of different BCG strains are used as vaccines, which have some other minor genetic differences (Table 34.5), although there is no evidence that these affect the protective efficacy of BCG. BCG was first used as a vaccine in 1921!
Table 34.5 Genetic deletions in different strains of BCG vaccine
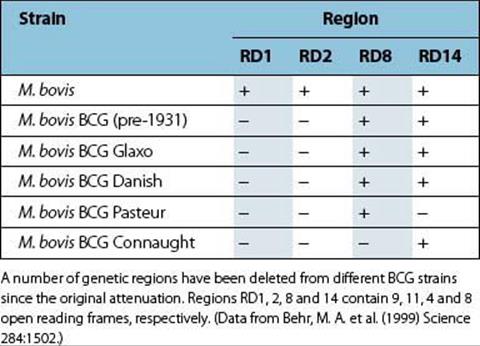
BCG is usually given to babies shortly after birth and is given to over 100 million children annually. It provides good (and very cost-effective) prevention of the disseminated forms of childhood tuberculosis, but variable protection against pulmonary TB in adults. For example, it induced good protection (> 80%) in trials in adolescents in the UK, but no protection in South India or Malawi. The reasons for this may include exposure to environmental mycobacteria that can induce a masking or a blocking effect on the immunity induced by BCG. When BCG is protective, this is associated with induction of a TH1 immune response – although simply measuring the IFNγ induced in response to mycobacterial antigens does not provide a correlate of protection. When induced, protective immunity lasts for 10–15 years and in one study was shown to last for over 50 years. In settings where BCG is protective, it may protect against infection as well as against disease. There is no evidence that revaccination is helpful. In children over 6 years of age, or in those known or likely to have been infected with M. tuberculosis, skin testing with M. tuberculosispurified protein derivative (Mantoux skin test) should be performed and BCG vaccination only given to those with a negative test result.
Because of the variable protection that BCG vaccination gives against tuberculosis in adults, the search is on for a new TB vaccine. Candidate vaccines in development include genetically modified BCGs, attenuated M. tuberculosis, viral vectors expressing key antigens of M. tuberculosis, and fusion proteins in adjuvant. A modified vaccinia virus Ankara expressing Ag85A is now in Phase IIb trials in children and in HIV-infected adults in Africa. This vaccine would be given as a boosting vaccine following BCG vaccination, and has been shown to boost the numbers of Ag85A-specific T cells secreting IFNγ (Fig. 34.10). Other promising vaccine candidates include genetically modified BCGs that express haemolysin or perfringolysin, that are designed to enhance activation of CD8 T cells through escape of antigens into the cytoplasm of the infected macrophage, and a fusion protein of Antigen85A with ESAT-6.
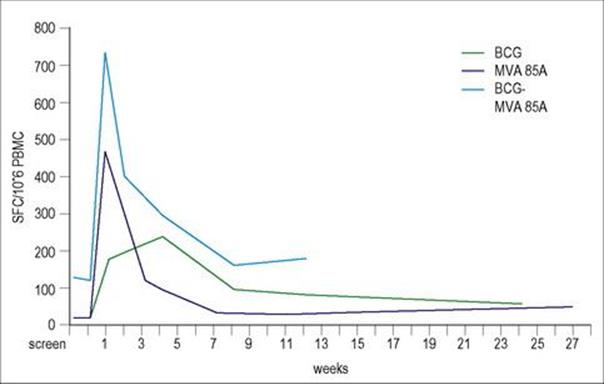
Figure 34.10 The prime boost strategy is being exploited in the design of new vaccines. A new candidate vaccine for TB uses modified vaccinia virus Ankara that expresses Antigen 85A (MVA85A) of M. tuberculosis to boost immunity in people previously vaccinated with BCG (BCG). The prime boost group of vaccinees (BCG-MVA85A) show the greatest numbers of spot-forming cells (SFC) making IFNγ in an ELISPOT assay in which peripheral blood mononuclear cells were stimulated with the Antigen 85 protein.
(Data from: McShane, H et al. (2004) Nature Medicine 10:1240–1244.)
Vaccines against hepatitis
The first vaccine for hepatitis B virus (HBV) consisted of the surface coat antigen of HBV purified from the plasma of virus carriers. This vaccine was protective, but required very careful purification and inactivation to ensure it was safe, and was expensive to produce. A recombinant hepatitis B vaccine was first licensed in the USA in 1986, and was the first vaccine produced using genetic engineering (Fig. 34.11). Recombinant HBV vaccines have an efficacy of 95% with immunity lasting at least 20 years after three doses of vaccine.
Figure 34.11 Electron micrograph of purified 22-nm hepatitis B surface antigens expressed in yeast cells.
(Courtesy of J.R. Pattison.)
Inactivated whole cell vaccines are available for hepatitis A. The virus is grown in human cells, purified, inactivated with formaldehyde and adsorbed onto alum. Again, these vaccines induce excellent immunity. However, as yet there is no vaccine available for hepatitis C.
Human papillomavirus (HPV)
New HPV vaccines have been introduced in the last decade, due to the association between HPV infection and cervical cancer. The first quadrivalent vaccine (Guardisil™), which induces immunity against four types of HPV, was licensed in 2006. This contains the L1 capsid protein of HPV from two oncogenic types of virus HPV16 and HPV18 as well as two non-oncogenic types, HBV6 and HPV11. It is made by recombinant DNA technology, expressed in yeast cells, and forms virus-like particles. This vaccine is being given to females aged 12–13, before they become sexually active, and can induce antibody responses in over 99.5% of vaccinees. The quadrivalent HPV vaccine is now also given to males aged 9–26 years of age in the USA to prevent genital warts as HPV6 and HPV11 cause approximately 90% of genital warts. A bivalent HPV vaccine containing L1 from HPV16 and HPV18 was approved for females aged between 10 and 25 years of age in the USA in 2009. This vaccine is cheaper, but most countries have opted for the quadrivalent vaccine, as it also prevents genital warts.
Rotavirus vaccine
Rotavirus is the cause of most serious gastrointestinal disease in infants. Trials of an earlier vaccine were stopped when it caused intussusception, a rare cause of bowel obstruction. Two new rotavirus vaccines were recommended for use by the WHO in 2009. Such viruses illustrate the difficult decisions to be made when there is a small risk of a vaccine-induced complication, but protection from disease would save many lives. Two new live vaccines are now in use: the RV5 oral vaccine (RotaTeq™) contains five reassortant rotaviruses developed from human and bovine parent strains, while the RV1 vaccine (RotarixTM) contains one live attenuated rotavirus strain. Trials of RV1 vaccine in 11 countries showed that three doses gave 98% protection against severe rotavirus gastroenteritis, while two doses of RV1 vaccine were shown to provide 85% protection against severe rotavirus gastroenteritis in Latin America.
Vaccines that are required for entry into particular countries, or for particular regions
The yellow fever vaccine is required for entry into certain countries. A vaccination certificate may be required for all those entering a particular country, or for individuals coming from a country where yellow fever is endemic. The certificate lasts for 10 years although immunity is probably life-long.
Pilgrims to Saudi Arabia for the Umrah or Haj may be required to show evidence of vaccination, for example, against meningitis (see above).
Travellers spending longer periods in areas of rural Asia, where Japanese Encephalitis (JE, a mosquito-transmitted flavivirus) is common, can be vaccinated with an inactivated JE virus vaccine. A cell-culture derived inactivated vaccine is now replacing an earlier mouse brain-derived inactivated vaccine. The cell-culture-derived vaccine requires two doses and is only given to those over 17 years of age; the mouse brain-derived vaccine requires three doses, and can be given to children and those under 17 years of age.
Vaccines for subgroups at high risk
Rabies vaccination is available for those exposed to rabies, or whose work or travel puts them at increased risk. Two types of vaccine are available: inactivated virus from cell cultures (from human diploid cells or chick embryo cell); intradermal vaccination may be more effective than intramuscular injection. Vaccine has also been prepared in human Vero cells. The cell culture-derived vaccines are considered safer than nerve tissue-based vaccines.
A vaccine has been produced for those working with Bacillus anthracis, such as laboratory or animal workers, or some military personnel. To ensure protection, five doses of vaccine are given and a yearly booster is necessary.
Complexity of vaccine schedules
An increasing number of vaccines are being given to infants – at a time when their immune system is not fully mature. However, studies have shown that pre-term babies can still be vaccinated safely at the right chronological age for vaccination. Table 34.6 gives an overview of vaccines being given to infants, children and adolescents in the USA and UK. The detailed schedules can be found on the WHO website (www.who.int).
Table 34.6 Examples of vaccination schedules in the UK and the USA
|
Vaccine |
UK |
USA |
|
Diphtheria, tetanus, acellular pertussis |
2, 3, 4 months |
2, 4, 6, 15–18 months, 4–6 years |
|
Inactivated polio vaccine |
2, 3.4 months |
2, 4, 6–18 months, 4–6 years |
|
Haemophilus influenzae type b |
2, 3, 4, 12–13 months |
2, 4, 6, 12–15 months |
|
Pneumococcal conjugate vaccine |
2, 4, 12–15 months |
2, 4, 6, 12–15 months |
|
Meningitis C |
3, 4, 12–13 months |
11–12 years, 16 years |
|
Measles, mumps and rubella |
12–13 months |
12–15 months, 4–6 years |
|
Hepatitis B |
Not used |
0, 1–3 months, 6–18 months |
|
Hepatitis A |
Not used |
12–13 months, 18–19 months |
|
Human papillomavirus* |
12–13 years |
11–12 years × 3** |
|
Varicella |
Not used |
12–15 months, 4–6 years |
|
Rotavirus |
Not used |
2, 4, (6) months*** |
|
Influenza |
From 6 months (pandemic vaccine only) |
From 6 months (seasonal vaccine) |
Note that the schedules and vaccines given differ. These indicative schedules are based on recommendations in April 2011; up-to-date- schedules can be found at http://www.nhs.uk/Planners/vaccinations/Pages/Vaccinationchecklist.aspx for the UK,http://www.cdc.gov/vaccines/recs/schedules/child-schedule.htm for the US and http://apps.who.int/immunization_monitoring/en/globalsummary/scheduleselect.cfm for all other countries.
* For human papillomavirus vaccine, the vaccine is routinely given to girls, but can be given to boys to prevent genital warts.
** Three doses given at 0, 1–2 months and 6 months schedule.
*** Different formulations of rotavirus vaccine require 2 or 3 doses.
It is important to ensure that all these vaccines do not interfere with each other, and thus reduce vaccine-induced immunity. It is therefore important to test for non-interference before a new vaccine is introduced.
There may be other factors that affect how well a vaccine works in the real world. Some studies have reported sex differences in vaccine-induced immunity, and seasonal effects, so some vaccines may not induce equivalent immunity in all settings. Vaccination is a very powerful public health tool, but not all infants and children will get their vaccines at the right ages or in the recommended order. Vaccines for developing countries therefore need to be tested in the populations most at risk, where other factors as well as infections such as malaria and intestinal helminths may modulate the immunity they induce.
Changes in demography means new vaccine strategies are needed
In many countries, the proportion of older individuals is increasing. With age, immunity can be lost, and in particular, T-cell immunity is weakened. Hospitalizations for infections such as pneumonia and influenza in older people place a burden on health systems. One strategy is to vaccinate older individuals against these diseases – but, due to the reduced efficiency of the immune system in old age, new vaccine strategies may be needed to overcome this immunosenescence, and the World Health Organization has identified the development of effective vaccines for the elderly as a research priority for 2010–2020.
New vaccines in development
Figure 34.12 shows that if effective vaccines were developed against HIV/AIDS, malaria and tuberculosis, many lives could be saved. The development of new vaccines against tuberculosis was covered above – but what about HIV and malaria?
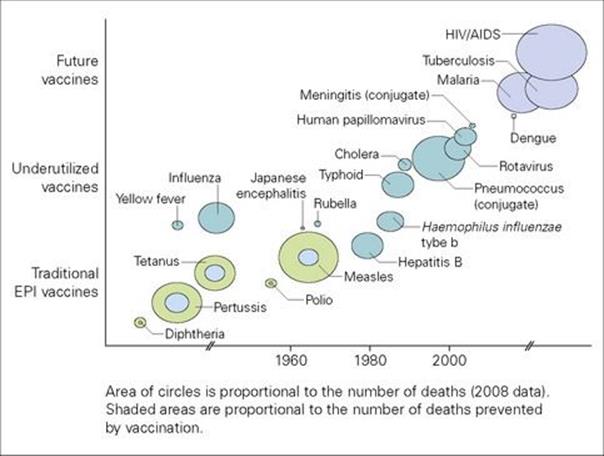
Figure 34.12 Deaths that could be averted by vaccination. Effective vaccines against HIV/AIDS, malaria and tuberculosis could save many lives, and a number of existing vaccines are underused. The area of the circles is proportional to the number of deaths in 2008, and shaded areas are proportional to the number of deaths prevented by vaccination.
(Redrawn from World Health Organization. The Initiative for Vaccine Research Strategic Plan 2010-2020. http://whqlibdoc.who.int/hq/2010/WHO_IVB_10.02_eng.pdf)
HIV vaccines
HIV has proved to be a real challenge in terms of vaccine development. Since 1987, over 30 vaccines have been tested in phase I or phase II trials, but despite all this effort, and two large-scale trials of vaccine based on the envelope gp120 protein, no effective vaccine is yet available. To date, vaccines based on recombinant Env gp120 proteins with adjuvant, HIV-1 DNA plasmids, antigens delivered by a variety of viral vectors, and varying prime-boost regimens have been tested. A trial in Thailand using a priming canary pox vaccine encoding the genes for HIV envelope, core and regulatory proteins and a boosting vaccine with recombinant gp120 showed that modest protection of 31.2% was achieved. However, a trial using a recombinant adenovirus 5 was not only not protective but seemed to increase the risk of infection in those who had antibodies to the adenovirus 5 strain before vaccination. Part of the problem is that the gp120 molecule mutates and circulating HIV viruses are highly variable; the killed virus is not sufficiently immunogenic to use as a vaccine, and the route of infection, mostly through the genital tract, means localized mucosal immunity is needed. This illustrates that despite huge advances in molecular biology and immunology, it can be difficult to design a protective vaccine. Some of the new strategies being investigated include novel antigen design strategies to induce neutralizing antibodies, and strategies to modify the body’s response to infection, for example to reduce the viral load in T cells in infected individuals or to stimulate the activity of the antibody-producing B cells.
Malaria
Malaria has been another tricky disease against which to develop an effective vaccine. Most efforts have been directed against antigens expressed during the sporozoite stage in the liver, or the blood stage. There are currently 11 such vaccines in clinical development. The RTS,S vaccine based on the liver-stage circumsporozoite protein has shown 30–50% efficacy in phase II trials – in challenge studies in the USA and in field trials in Africa (Fig. 34.13), and a phase III trial is underway in Africa. In one respect, it should be possible to develop a vaccine against disease, if not against infection, as most individuals living in endemic areas develop a way of remaining well despite sometimes having circulating parasites in their blood. On the other hand, immunity is never complete in these people, and those who move away from endemic areas quickly lose this immunity. It has been suggested that a vaccine may need to induce immunity against malaria antigens that are not normally immunogenic and therefore may be under less immune pressure to change. A transmission-blocking vaccine against the sexual forms of malaria in the mosquito could also help reduce transmission.
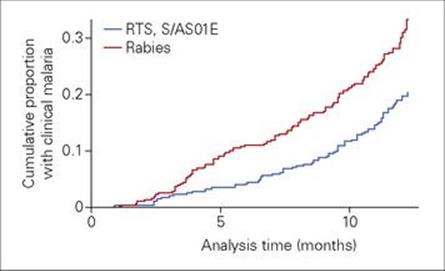
Figure 34.13 A new candidate vaccine for malaria that uses parts of the circumsporozoite protein fused to the hepatitis B surface antigen reduces the prevalence of malaria infection in young African children. Children given three doses of the RTS,S vaccine had a longer delay before they developed clinical malaria infection compared with controls given rabies vaccine.
(Data from Olutu, A. et al. (2011) Lancet Infect Dis 11: 102–109.)
Vaccines for neglected tropical diseases
Some infections such as leishmaniasis, leprosy, and helminth infections are described as neglected tropical diseases, neglected while most emphasis is put on HIV, malaria and TB. Schistosomiasis, leishmaniasis and trachoma are examples of neglected tropical diseases where there is no vaccine available. A vaccine against hookworm based on two key antigens is currently being developed. Leprosy also has no vaccine, but luckily the BCG vaccine has been shown to provide partial immunity to leprosy – hopefully any new TB vaccine will do even better.
New delivery systems and technologies for future vaccines
Adenoviruses are being tested as vaccine vectors, as they induce good CD8 T-cell responses. Some adenoviruses are not ideal vaccine vectors as too many individuals already have antibodies to them, which may reduce the efficacy of the vaccine. For example, although only 20% of individuals in the Netherlands have antibodies to type 5 adenovirus, this rises to 80% in sub-Saharan Africa, so some new vaccine trials are using the Ad35 strain instead, as the seroreactivity to Ad35 is lower in Africa. Delivery by MVA (see Fig. 34.10) or adenovirus is being used in the design of new vaccines for TB.
Genetic engineering can be used to make more effective vaccines. Viral recombinant vaccines are being developed as new RSV vaccines – using parainfluenza virus expressing key RSV proteins. Codon optimization can also be used, for example, for poliovirus, where reversion to virulence can be reduced by altering the codon usage.
Virus-like particles can be made that express the key viral proteins, yet are replication deficient. The latest papillomavirus vaccines are virus-like particles made from recombinant HPV coat proteins. This approach is being used for blue tongue virus vaccine for sheep, and is being considered for flu.
DNA vaccines were thought to hold great promise – but so far have not been licensed for use. They are good at priming the immune system and can induce good memory responses and TH1 responses in immunologically naive recipients.
Gentically modified or transgenic plants can be used to produce immunogens, including glycosylated proteins, and even virus-like particles. For example, the recombinant HBV vaccine could be produced in plants rather than in yeast.
New routes of vaccination
The oral polio vaccine was not the first vaccine to be given orally – the BCG vaccine was originally given by mouth. Dissolvable tablets or wafers may be used under the tongue in future. Some new work is even investigating expressing vaccine antigens in edible fruit or vegetables, such as tomatoes or lettuce!
If protection is needed in the mucosal-associated lymphoid tissues, then to prime cells in this region is very sensible, and nasal sprays can be used, as in one current formulation of the seasonal flu vaccine in the USA. Another approach is to use skin patches – these deliver the vaccine antigens through the transcutaneous route. Vaccines of the future may even use nanoparticles, or be injected using dissolving microneedles, said to be relatively painless. This is clearly an area where molecular science and technological developments can make a real impact.
The Decade of Vaccines
The Bill and Melinda Gates Foundation called in 2010 for this to be the Decade of Vaccines. As a result, the World Health Organization (WHO), UNICEF, the National Institute of Allergy and Infectious Diseases (NIAID) and the Bill & Melinda Gates Foundation are collaborating to produce a Global Vaccine Action Plan. The technology to make many new vaccines is now available, but many challenges remain including how to shorten the gap between vaccine discovery and licensing, how to provide equitable access to vaccines at an affordable price, how to overcome ‘vaccine hesitancy’ within communities, and how to strengthen health systems so that the vaccines we have, and that work, can be fully utilized.
![]()
 Key Facts
Key Facts
• Vaccination aims to prime the adaptive immune system to the antigens of a particular microbe so that a first infection induces a secondary immune response.
• Vaccines can use live attenuated organisms, killed whole organisms, subcellular fractions or antigens produced artificially by gene cloning or chemical synthesis.
• In general, live vaccines are more effective than other types, but carry the risk of reverting to virulence or inducing disease in immunocompromised patients.
• The details of vaccine choice, route, dose and risks have to be considered for each disease individually.
• Overall, vaccination is a very effective public health tool, but many challenges remain, including the effective implementation of existing vaccines worldwide and the design of new vaccines against those infections for which they are not yet available.
![]()