CONCEPTS IN BIOLOGY
PART II. CORNERSTONES: CHEMISTRY, CELLS, AND METABOLISM
7. Biochemical Pathways-Photosynthesis
7.3. The Metabolic Pathways of Photosynthesis
It is a good idea to begin with the simplest description and add layers of understanding as you go to additional levels. Therefore, this discussion of photosynthesis is divided into two levels:
1. a fundamental description, and
2. a detailed description.
Ask your instructor which level is required for your course of study.
Fundamental Description
Light-Capturing Events
Light energy is used to drive photosynthesis during the lightcapturing events. Visible light is a combination of many different wavelengths of light, seen as different colors. Some of these colors appear when white light is separated into its colors to form a rainbow. The colors of the electromagnetic spectrum that provide the energy for photosynthesis are correlated with different kinds of light-energy-absorbing pigments. The green chlorophylls are the most familiar and abundant. There are several types of this pigment. The two most common types are chlorophyll a and chlorophyll b. Both absorb strongly in the red and blue portions of the electromagnetic spectrum, although in slightly different portions of the spectrum (figure 7.4).

Chlorophylls reflect green light. That is why we see chlorophyll-containing plants as predominantly green. Other pigments common in plants are called accessory pigments. These include the carotenoids (yellow, red, and orange). They absorb mostly blue and blue-green light while reflecting the oranges and yellows. The presence of these pigments is generally masked by the presence of chlorophyll, but in the fall, when chlorophyll disintegrates, the reds, oranges, and yellows show through. Accessory pigments are also responsible for the brilliant colors of vegetables, such as carrots, tomatoes, eggplant, and peppers. Photosynthetic bacteria and various species of algae have other kinds of accessory pigments not found in plants. Having a combination of different pigments, each of which absorbs a portion of the light spectrum hitting it, allows the organism to capture much of the visible light that falls on it.
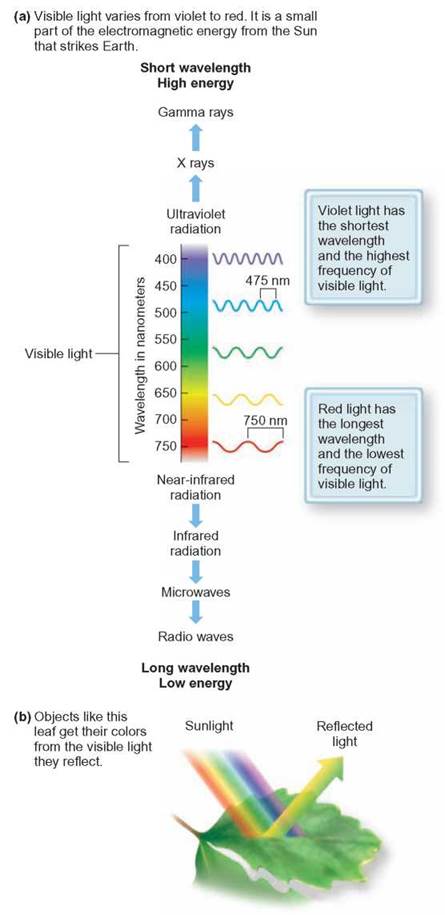
FIGURE 7.4. The Electromagnetic Spectrum, Visible Light, and Chlorophyll
Light is a form of electromagnetic energy that can be thought of as occurring in waves. Chlorophyll absorbs light most strongly in the blue and red portion of the electromagnetic spectrum but poorly in the green portions. The shorter the wavelength, the more energy it contains. Humans are capable of seeing only waves that are between about 400 and 740 nanometers (nm) long.
Any cell with chloroplasts can carry on photosynthesis. However, in most plants, leaves are specialized for photosynthesis and contain cells that have high numbers of chloroplasts (figure 7.5).
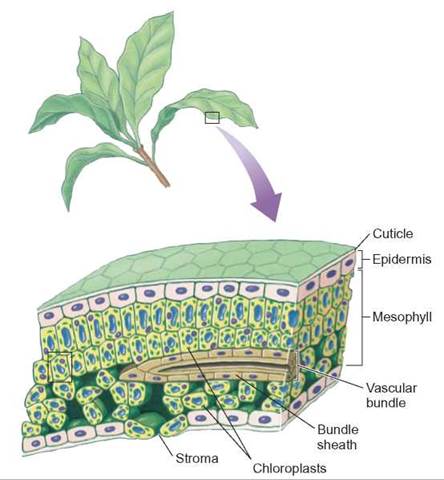
FIGURE 7.5. Photosynthesis and the Structure of a Plant Leaf
Plant leaves are composed of layers of cells that contain chloroplasts, which contain chlorophyll.
Chloroplasts are membrane-enclosed organelles that contain many thin, flattened sacs that contain chlorophyll. These chlorophyll-containing sacs are called thylakoids and a number of these thylakoids stacked together is known as a granum. In addition to chlorophyll, the thylakoids contain accessory pigments, electron-transport molecules, and enzymes. Recall that the fluid-filled spaces between the grana are called the stroma of the chloroplast. The structure of the chloroplast is directly related to both the light-capturing and the energy-conversion steps of photosynthesis. In the light-capturing events, the pigments (e.g., chlorophyll), which are embedded in the membranes of the thylakoids, capture light energy and some of the electrons of pigments become excited. The chlorophylls and other pigments involved in trapping sunlight energy and storing it are arranged into clusters called photosystems. By clustering the pigments, photosystems serve as energy-gathering, or energyconcentrating, mechanisms that allow light to be collected more efficiently and excite electrons to higher energy levels.
A Fundamental Summary of Light-Capturing Events
photons of light energy → excited electrons from chlorophyll
Light-Dependent Reactions
The light-dependent reactions of photosynthesis also take place in the thylakoid membranes inside the chloroplast. The excited electrons from the light-capturing events are passed to protein molecules in the thylakoid membrane. The electrons are passed through a series of electron-transport steps, which result in protons being pumped into the cavity of the thylakoid. When the protons pass back out through the membrane to the outside of the thylakoid, ATP is produced. This is very similar to the reactions that happen in the electron-transport system (ETS) of aerobic cellular respiration. In addition, the chlorophyll that just lost its electrons to the chloroplast’s electron-transport system regains electrons from water molecules. This results in the production of hydrogen ions, electrons, and oxygen gas. The next light-capturing event will excite this new electron and send it along the electron-transport system. As electrons finish moving through the electron-transport system, the coenzyme NADP+ picks up the electrons and is reduced to NADPH. The hydrogen ions attach because, when NADP+ accepts electrons, it becomes negatively charged (NADP-). The positively charged H+ are attracted to the negatively charged NADP-. The oxygen remaining from the splitting of water molecules is released into the atmosphere, or it can be used by the cell in aerobic cellular respiration, which takes place in the mitochondria of plant cells. The ATP and NADPH molecules move from the grana, where the light-dependent reactions take place, to the stroma, where the light-independent reactions take place.
A Fundamental Summary of the Light-Dependent Reactions
excited electrons + H2O + ADP + NADP+ → ATP + NADPH + O2
Light-Independent Reactions
The ATP and NADPH provide energy, electrons and hydrogens needed to build large, organic molecules. The light-independent reactions are a series of oxidation-reduction reactions, which combine hydrogen from water (carried by NADPH) with carbon dioxide from the atmosphere to form simple organic molecules, such as sugar. As CO2 diffuses into the chloroplasts, the enzyme Ribulose-1,5-bisphosphate carboxylase oxygenase (RuBisCO) speeds the combining of the CO2 with an already present 5-carbon sugar, ribulose. NADPH then donates its hydrogens and electrons to complete the reduction of the molecule. The resulting 6-carbon molecule is immediately split into two 3-carbon molecules of glyceraldehyde-3-phosphate. Some of the glyceraldehyde-3-phosphate molecules are converted through another series of reactions into ribulose. Thus, these reactions constitute a cycle, in which carbon dioxide and hydrogens are added and glyceraldehyde-3-phosphate and the original 5-carbon ribulose are produced. The plant can use surplus glyceraldehyde-3-phosphate for the synthesis of glucose. The plant can also use glyceraldehyde-3-phosphate to construct a wide variety of other organic molecules (e.g., proteins, nucleic acids), provided there are a few additional raw materials, such as minerals and nitrogen-containing molecules (figure 7.6).
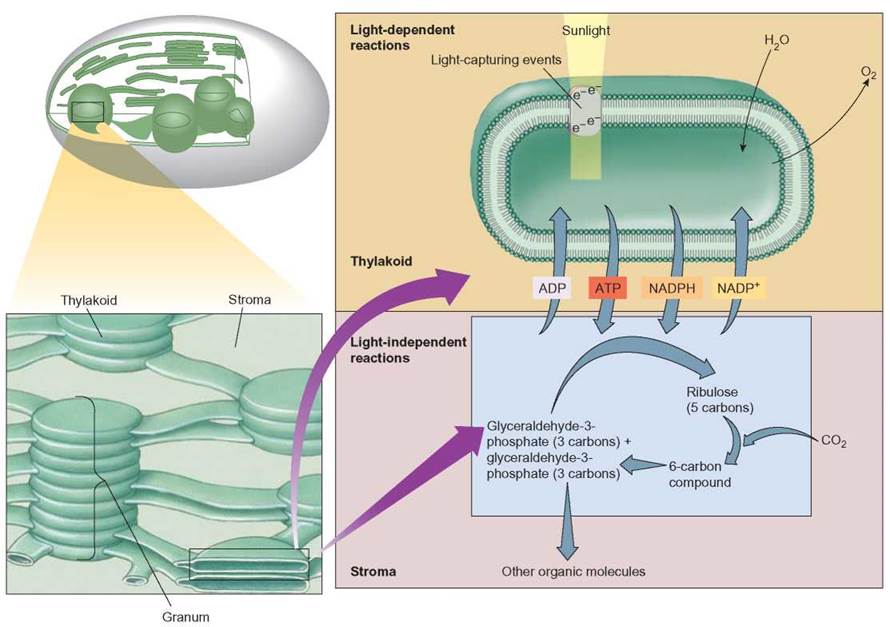
FIGURE 7.6. Photosynthesis: Fundamental Description
The process of photosynthesis involves light-capturing events by chlorophyll and other pigments. The excited electrons are used in the light-dependent reactions to split water, releasing hydrogens and oxygen. The hydrogens are picked up by NADP+ to form NADPH and the oxygen is released. Excited electrons are also used to produce ATP. The ATP and NADPH leave the thylakoid and enter the stroma of the chloroplast, where they are used in the light-independent reactions to incorporate carbon dioxide into organic molecules. During the light-independent reactions, carbon dioxide is added to a 5-carbon ribulose molecule to form a 6-carbon compound, which splits into glyceraldehyde-3-phosphate. Some of the glyceraldehyde-3-phosphate is used to regenerate ribulose and some is used to make other organic molecules. The ADP and NADP+ released from the light-independent reactions stage return to the thylakoid to be used in the synthesis of ATP and NADPH again. Therefore, each stage is dependent on the other.
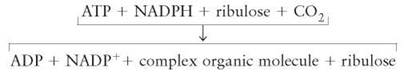
A Fundamental Summary of the Light-Independent Reactions
Detailed Description
Light-Capturing Events
The energy of light comes in discrete packages, called photons. Photons of light having different wavelengths have different amounts of energy. A photon of red light has a different amount of energy than a photon of blue light. Pigments of different kinds are able to absorb photons of certain wavelengths of light. Chlorophyll absorbs red and blue light best and reflects green light. When a chlorophyll molecule is struck by and absorbs a photon of the correct wavelength, its electrons become excited to a higher energy level. This electron is replaced when chlorophyll takes an electron from a water molecule. The excited electron goes on to form ATP. The reactions that result in the production of ATP and the splitting of water take place in the thylakoids of chloroplasts. There are many different molecules involved, and most are embedded in the membrane of the thylakoid. The various molecules involved in these reactions are referred to as photosystems. A photosystem is composed of (1) an antenna complex, (2) a reaction center, and (3) other enzymes necessary to store the light energy as ATP and NADPH.
The antenna complex is a network of hundreds of chlorophyll and accessory pigment molecules, whose role is to capture photons of light energy and transfer the energy to a specialized portion of the photosystem known as the reaction center. When light shines on the antenna complex and strikes a chlorophyll molecule, an electron becomes excited. The energy of the excited electron is passed from one pigment to another through the antenna complex network. This series of excitations continues until the combined energies from several excitations are transferred to the reaction center, which consists of a complex of chlorophyll a and protein molecules. An electron is excited and passed to a primary electron acceptor molecule, oxidizing the chlorophyll and reducing the acceptor. Ultimately, the oxidized chlorophyll then has its electron replaced with another electron from a different electron donor. Exactly where this replacement electron comes from is the basis on which two different photosystems have been identified—photosystem I and photosystem II, which will be discussed in the next section.
Summary of Detailed Description of the Light-Capturing Reactions
1. They take place in the thylakoids of the chloroplast.
2. Chlorophyll and other pigments of the antenna complex capture light energy and produce excited electrons.
3. The energy is transferred to the reaction center.
4. Excited electrons from the reaction center are transferred to a primary electron acceptor molecule.
Light-Dependent Reactions
Both photosystems I and II have antenna complexes and reaction centers and provide excited electrons to primary electron acceptors. However, each has slightly different enzymes and other proteins associated with it, so each does a slightly different job. In actuality, photosystem II occurs first and feeds its excited electrons to photosystem I (figure 7.7). One special feature of photosystem II is that there is an enzyme in the thylakoid membrane responsible for splitting water molecules (H2O → 2H + O). The oxygen is released as O2 and the electrons of the hydrogens are used to replace the electrons that had been lost by the chlorophyll. The remaining hydrogen ions (protons) are released to participate in other reactions. Thus, in a sense, the light energy trapped by the antenna complex is used to split water into H and O. The excited electrons from photosystem II are sent through a series of electron-transport reactions, in which they give up some of their energy. This is similar to the electron-transport system of aerobic cellular respiration. After passing through the electron-transport system, the electrons are accepted by chlorophyll molecules in photosystem I. While the electron-transport activity is happening, protons are pumped from the stroma into the space inside the thylakoid. Eventually, these protons move back across the membrane. When they do, ATPase is used to produce ATP (ADP is phosphorylated to produce ATP). Thus, a second result of this process is that the energy of sunlight has been used to produce ATP.
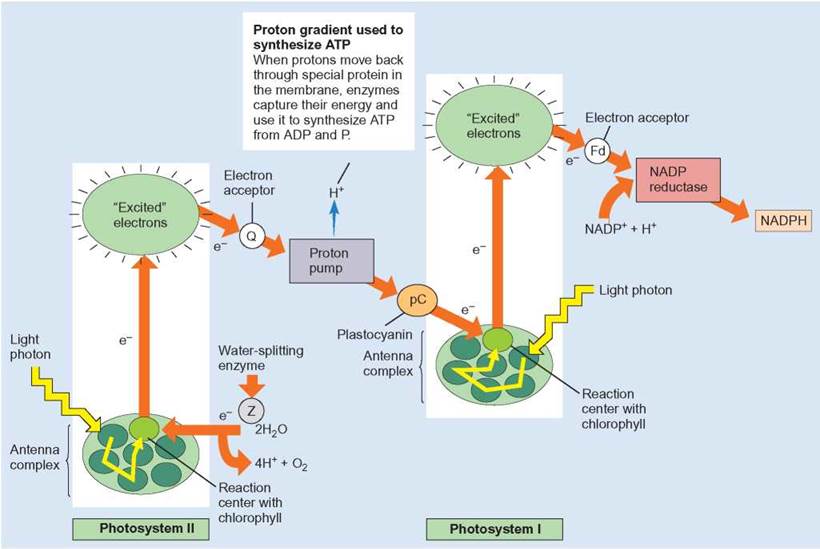
FIGURE 7.7. Photosystems II and I and How They Interact: Detailed Description
Although light energy strikes and is absorbed by both photosystem II and I, what happens and how they interconnect are not the same. Notice that the electrons released from photosystem II end up in the chlorophyll molecules of photosystem I. The electrons that replace those “excited” out of the reaction center in photosystem II come from water.
The connection between photosystem II and photosystem I involves the transfer of electrons from photosystem II to photosystem I. These electrons are important because photons (from sunlight) are exciting electrons in the reaction center of photosystem I and the electrons from photosystem II replace those lost from photosystem I.
In photosystem I, light is trapped and the energy is absorbed in the same manner as in photosystem II. However, this system does not have the enzyme involved in splitting water; therefore, no O2is released from photosystem I. The high-energy electrons leaving the reaction center of photosystem I make their way through a different series of oxidation-reduction reactions. During these reactions, the electrons are picked up by NADP+, which is reduced to NADPH (review figure 7.7). Thus, the primary result of photosystem I is the production of NADPH.
Summary of Detailed Description of the Light-Dependent Reactions of Photosynthesis
1. They take place in the thylakoids of the chloroplast.
2. Excited electrons from photosystem II are passed through an electron-transport chain and ultimately enter photosystem I.
3. The electron-transport system is used to establish a proton gradient, which produces ATP.
4. Excited electrons from photosystem I are transferred to NADP+ to form NADPH.
5. In photosystem II, an enzyme splits water into hydrogen and oxygen. The oxygen is released as O2.
6. Electrons from the hydrogen of water replace the electrons lost by chlorophyll in photosystem II.
Light-Independent Reactions
The light-independent reactions take place within the stroma of the chloroplast. The materials needed for the light-independent reactions are ATP, NADPH, CO2, and a 5-carbon starter molecule called ribulose. The first two ingredients (ATP and NADPH) are made available from the light-dependent reactions, photosystems II and I. The carbon dioxide molecules come from the atmosphere, and the ribulose starter molecule is already present in the stroma of the chloroplast from previous reactions.
Carbon dioxide is said to undergo carbon fixation through the Calvin cycle (named after its discoverer, Melvin Calvin). In the Calvin cycle, ATP and NADPH from the light-dependent reactions are used, along with carbon dioxide, to synthesize larger, organic molecules. As with most metabolic pathways, the synthesis of organic molecules during the light-independent reactions requires the activity of several enzymes to facilitate the many steps in the process. The fixation of carbon begins with carbon dioxide combining with the 5-carbon molecule ribulose to form an unstable 6-carbon molecule. This reaction is carried out by the enzyme Ribulose-1,5-bisphosphate carboxylase oxygenase (RuBisCO), reportedly the most abundant enzyme on the planet. The newly formed 6-carbon molecule immediately breaks down into two 3-carbon molecules, each of which then undergoes a series of reactions involving a transfer of energy from ATP and a transfer of hydrogen from NADPH. The result of this series of reactions is two glyceraldehyde-3-phosphate molecules. Because glyceraldehyde-3-phosphate contains 3 carbons and is formed as the first stable compound in this type of photosynthesis, this is sometimes referred to as the C3 photosynthetic pathway. Some of the glyceraldehyde-3-phosphate is used to synthesize glucose and other organic molecules, and some is used to regenerate the 5-carbon ribulose molecule, so this pathway is a cycle (figure 7.8). Outlooks 7.1 describes some other forms of photosynthesis that do not use the C3 pathway.
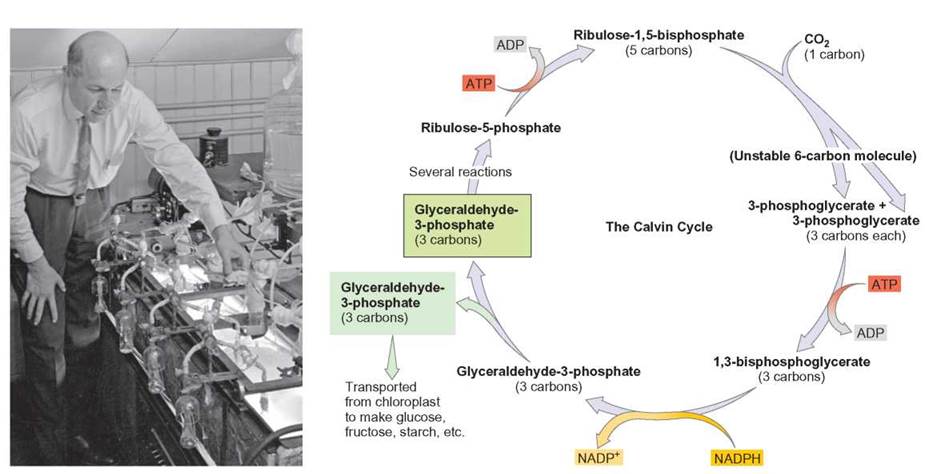
FIGURE 7.8. The Calvin Cycle: Detailed Description
During the Calvin cycle, ATP and NADPH from the light-dependent reactions are used to attach CO2 to the 5-carbon ribulose molecule.
The 6-carbon molecule formed immediately breaks down into two 3-carbon molecules. Some of the glyceraldehyde-3-phosphate formed is used to produce glucose and other, more complex organic molecules. In order to accumulate enough carbon to make a new glucose molecule, the cycle must turn six times. The remaining glyceraldehyde-3-phosphate is used to regenerate the 5-carbon ribulose to start the process again.
Summary of Detailed Description of the Reactions of the Light-Independent Events
1. They take place in the stroma of chloroplasts:
2. ATP and NADPH from the light-dependent reactions leave the grana and enter the stroma.
3. The energy of ATP is used in the Calvin cycle to combine carbon dioxide to a 5-carbon starter molecule (ribulose) to form a 6-carbon molecule.
4. The 6-carbon molecule immediately divides into two 3-carbon molecules of glyceraldehyde-3-phosphate.
5. Hydrogens from NADPH are transferred to molecules in the Calvin cycle.
6. The 5-carbon ribulose is regenerated.
7. ADP and NADP+ are returned to the light-dependent reactions.
OUTLOOKS 7.1
The Evolution of Photosynthesis
It is amazing that the processes of photosynthesis in prokaryotes and eukaryotes are so similar. The evolution of photosynthesis goes back over 3 billion years, when all life on Earth was prokaryotic and occurred in organisms that were aquatic. (There were no terrestrial organisms at the time.) Today, some bacteria perform a kind of photosynthesis that does not result in the release of oxygen. In general, these bacteria produce ATP but do not break down water to produce oxygen. Perhaps these are the descendents of the first organisms to carry out a photosynthetic process, and oxygen-releasing photosynthesis developed from these earlier forms of photosynthesis.
Evidence from the fossil record shows that, beginning approximately 2.4 billion years ago, oxygen was present in the atmosphere. Eukaryotic organisms had not yet developed, so the organisms responsible for producing this oxygen would have been prokaryotic. Today, many kinds of cyanobacteria perform photosynthesis in essentially the same way as plants, although they use a somewhat different kind of chlorophyll. As a matter of fact, it is assumed that the chloroplasts of eukaryotes are evolved from photosynthetic bacteria. Initially, the first eukaryotes to perform photosynthesis would have been various kinds of algae. Today, certain kinds of algae (red algae, brown algae, green algae) have specific kinds of chlorophylls and other accessory pigments different from the others. Because the group known as the green algae has the same chlorophylls as plants, it is assumed that plants are derived from this aquatic group.
The evolution of photosynthesis did not stop once plants came on the scene, however. Most plants perform photosynthesis in the manner described in this chapter. Light energy is used to generate ATP and NADPH, which are used in the Calvin cycle to incorporate carbon dioxide into glyceraldehyde-3-phosphate. Because the primary product of this form of photosynthesis is a 3-carbon molecule of glyceraldehyde-3-phosphate, it is often called C3 photosynthesis. Among plants, there are two interesting variations of photosynthesis, which use the same basic process but add interesting twists.
C4 photosynthesis is common in plants like grasses, such as corn (maize), crabgrass, and sugarcane that are typically subjected to high light levels. In these plants, carbon dioxide does not directly enter the Calvin cycle. Instead, the fixation of carbon is carried out in two steps, and two kinds of cells participate. It appears that this adaptation allows plants to trap carbon dioxide more efficiently from the atmosphere under high light conditions. Specialized cells in the leaf capture carbon dioxide and convert a 3-carbon compound to a 4-carbon compound. This 4-carbon compound then releases the carbon dioxide to other cells, which use it in the normal Calvin cycle typical of the light-independent reactions. Because a 4-carbon molecule is formed to "store" carbon, this process is known as C4 photosynthesis.
Another variation of photosynthesis is known as Crassulacean acid metabolism (CAM), because this mechanism was first discovered in members of the plant family, Crassulaceae. (A common example, Crassula, is known as the jade plant.) CAM photosynthesis is a modification of the basic process of photosynthesis that allows photosynthesis to occur in arid environments while reducing the potential for water loss. In order for plants to take up carbon dioxide, small holes in the leaves (stomata) must be open to allow carbon dioxide to enter. However, relative humidity is low during the day and plants would tend to lose water if their stomates were open. CAM photosynthesis works as follows: During the night, the stomates open and carbon dioxide can enter the leaf. The chloroplasts trap the carbon dioxide by binding it to an organic molecule, similar to what happens in C4 plants. The next morning, when it is light (and drier), the stomates close. During the day, the chloroplasts can capture light and run the light-dependent reactions. They then use the carbon stored the previous night to do the light-independent reactions.



Glyceraldehyde-3-Phosphate: The Product of Photosynthesis
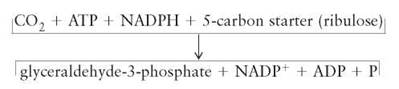
The 3-carbon glyceraldehyde-3-phosphate is the actual product of the process of photosynthesis. However, many textbooks show the generalized equation for photosynthesis as
![]()
making it appear as if a 6-carbon sugar (hexose) were the end product. The reason a hexose (C6H12O6) is usually listed as the end product is simply because, in the past, the simple sugars were easier to detect than was glyceraldehyde-3-phosphate.
Several things can happen to glyceraldehyde-3-phosphate. If a plant goes through photosynthesis and produces 12 glyceraldehyde-3-phosphates, 10 of the 12 are rearranged by a series of complex chemical reactions to regenerate the 5-carbon ribulose needed to operate the light-independent reactions stage. The other two glyceraldehyde-3-phosphates can be considered profit from the process. The excess glyceraldehyde-3-phosphate molecules are frequently changed into a hexose. So, the scientists who first examined photosynthesis chemically thought that sugar was the end product. It was only later that they realized that glyceraldehyde-3-phosphate is the true end product of photosynthesis.
Cells can do a number of things with glyceraldehyde-3-phosphate, in addition to manufacturing hexose (figure 7.9). Many other organic molecules can be constructed using glyceraldehyde-3-phosphate. Glyceraldehyde-3-phosphate can be converted to glucose molecules, which can be combined to form complex carbohydrates, such as starch for energy storage or cellulose for cell wall construction. In addition, other simple sugars can be used as building blocks for ATP, RNA, DNA, and other carbohydrate-containing materials.
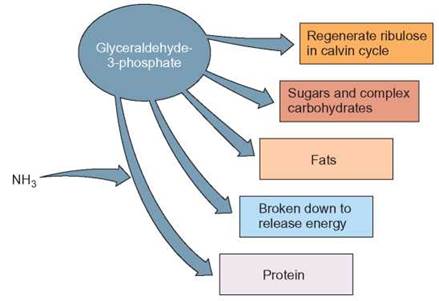
FIGURE 7.9. Uses for Glyceraldehyde-3-Phosphate
The glyceraldehyde-3-phosphate that is produced as the end product of photosynthesis has a variety of uses. The plant cell can make simple sugars, complex carbohydrates, or even the original 5-carbon starter from it. The glyceraldehyde-3-phosphate can also serve as an ingredient of lipids and amino acids (proteins). In addition, it is a major source of metabolic energy provided from aerobic respiration in the mitochondria of plant cells.
The cell can also convert the glyceraldehyde-3- phosphate into lipids, such as oils for storage, phospholipids for cell membranes, or steroids for cell membranes. The glyceraldehyde-3-phosphate can serve as the carbon skeleton for the construction of the amino acids needed to form proteins. Almost any molecule that a green plant can manufacture begins with this glyceraldehyde-3-phosphate molecule. Finally, glyceraldehyde-3-phosphate can be broken down during cellular respiration. Cellular respiration releases the chemical-bond energy from glyceraldehyde-3-phosphate and other organic molecules and converts it into the energy of ATP. This conversion of chemical-bond energy enables the plant cell and the cells of all organisms to do things that require energy, such as grow and move materials (Outlooks 7.2).
OUTLOOKS 7.2
Even More Ways to Photosynthesize
Having gone through the information on photosynthesis, you might have thought that this was the only way for this biochemical pathway to take place. However, there are many prokaryotes capable of carrying out photosynthesis using alternative pathways. These Bacteria and Archaea have light-capturing pigments, but they are not the same as plant chlorophylls or the accessory pigments. The range of light absorption differs, allowing many of these Bacteria and Archaea to live in places unfriendly to plants. Some forms of photosynthetic bacteria do not release oxygen, but rather release other by-products such as H2, H2S, S, or organic compounds. Table 7.1 compares some of the most important differences between eukaryotic and prokaryotic photosynthesis.
TABLE 7.1. Different Types of Photosynthesis
|
Property |
Eukaryotic |
Prokaryotic—Cyanobacteria |
Prokaryotic—Green and Purple Bacteria |
|
Photosystem pigments |
Chlorophyll a, b, and accessory pigments |
Chlorophyll a and phycocyanin (blue- green pigment) |
Combinations of bacteriochlorophylls a, b, c, d, or e absorb different wavelengths of light and some absorb infrared light. |
|
Thylakoid system |
Present |
Present |
Absent—pigments are found in vesicles called chlorosomes, or they are simply attached to plasma membrane. |
|
Photosystem II |
Present |
Present |
Absent |
|
Source of electrons |
H2O |
H2O |
H2, H2S, S, or a variety of organic molecules |
|
O2 production pattern |
Oxygenic— release O2 |
Oxygenic |
Anoxygenic—do not release O2 May release S, other organic compounds other than that used as the source of electrons |
|
Primary products of |
ATP + NADPH |
ATP + NADPH |
ATP |
|
energy conversion Carbon source Example |
CO2 Maple tree—Acer |
CO2 Anabaena Ocillatoria Nostoc |
Organic and/or CO2 Green sulfur bacterium—Chlorobium Green nonsulfur bacterium—Chloroflexus Purple sulfur bacterium—Chromatium Purple nonsulfur bacterium—Rhodospirillum |
7.3. CONCEPT REVIEW
5. How do photosystem I and photosystem II differ in the kinds of reactions that take place?
6. What does an antenna complex do? How is it related to the reaction center?
7. What role is played by the compound Ribulose-1,5-bisphosphate carboxylase oxygenase (RuBisCo)?
8. What role is played by the compound glyceraldehyde-3-phosphate?
9. Describe how photosystem II interacts with photosystem I.
10. What is the value of a plant to have more than one kind of photosynthetic pigment?