THE LIVING WORLD
Unit Five. Evolution of Animal Life
19. Evolution of the Animal Phyla
19.11. Protostomes and Deuterostomes
All the animals we have met so far have essentially the same kind of embryonic development. Cell divisions of the fertilized egg produce a hollow ball of cells, a blastula, which indents to form a two-layer-thick ball with a blastopore opening to the outside. In mollusks, annelids, and arthropods, the mouth (stoma) develops from or near the blastopore. An animal whose mouth develops in this way is called a protostome (figure 19.26, top). If such an animal has a distinct anus or anal pore, it develops later in another region of the embryo.
A second distinct pattern of embryological development occurs in the echinoderms and the chordates. In these animals, the anus forms from or near the blastopore, and the mouth forms subsequently on another part of the blastula. This group of phyla consists of animals that are called the deuterostomes (figure 19.26, bottom).
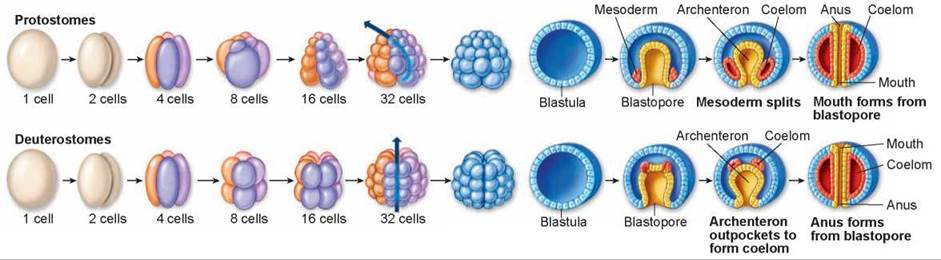
Figure 19.26. Embryonic development in protostomes and deuterostomes.
Cleavage of the egg produces a hollow ball of cells called the blastula. Invagination, or infolding, of the blastula produces the blastopore. In protostomes, embryonic cells cleave in a spiral pattern and become tightly packed. The blastopore becomes the animal's mouth, and the coelom originates from a mesodermal split. In deuterostomes, embryonic cells cleave radially and form a loosely packed array. The blastopore becomes the animal's anus, and the mouth develops at the other end. The coelom originates from an evagination, or outpouching, of the archenteron in deuterostomes.
Deuterostomes represent a revolution in embryonic development. In addition to the fate of the blastopore, deutero-stomes differ from protostomes in three other features:
1. The progressive division of cells during embryonic growth is called cleavage. The cleavage pattern relative to the embryo’s polar axis determines how the cells array. In nearly all protostomes, each new cell buds off at an angle oblique to the polar axis. As a result, a new cell nestles into the space between the older ones in a closely packed array (see the 16-cell stage in the upper row of cells). This pattern is called spiral cleavage because a line drawn through a sequence of dividing cells spirals outward from the polar axis, (indicated by the curving blue arrow at the 32-cell stage).
In deuterostomes, the cells divide parallel to and at right angles to the polar axis. As a result, the pairs of cells from each division are positioned directly above and below one another (see the 16-cell stage in the lower row of cells); this process gives rise to a loosely packed array of cells. This pattern is called radial cleavage because a line drawn through a sequence of dividing cells describes a radius outward from the polar axis (indicated by the straight blue arrow at the 32-cell stage).
2. In protostomes, the developmental fate of each cell in the embryo is fixed when that cell first appears. Even at the four-celled stage, each cell is different, containing different chemical developmental signals and no one cell, if separated from the others, can develop into a complete animal. In deuterostomes, on the other hand, the first cleavage divisions of the fertilized embryo produce identical daughter cells, and any single cell, if separated, can develop into a complete organism.
3. In all coelomates, the coelom originates from mesoderm. In protostomes, this occurs simply and directly: The mesoderm cells simply move away from one another as the coelomic cavity expands within the mesoderm. However, in deuterostomes, the coelom is normally produced by an evagination of the archenteron—the main cavity within the gastrula, also called the primitive gut. This cavity, lined with endoderm, opens to the outside via the blastopore and eventually becomes the gut cavity. The evaginating cells give rise to the mesodermal cells, and the mesoderm expands to form the coelom.
Key Learning Outcome 19.11. In protostomes, the egg cleaves spirally, and the blastopore becomes the mouth. In deuterostomes, the egg cleaves radially, and the blastopore becomes the animal's anus.
A Closer Look
Diversity Is Only Skin Deep
Perhaps the most important lesson to emerge from the study of animal diversity is not the incredible variety of animal forms, from worms and spiders to sharks and antelopes, but rather their deep similarities. The body plans of all animals are assembled along a similar path, as if from the same basic blueprint. The same genes play critical roles throughout the animal kingdom, small changes in how they are activated leading to very different body forms.
The molecular mechanisms used to orchestrate development are thought to have evolved very early in the history of multicellular life. Animals utilize transcription factors, like those discussed on page 247, to turn on or off particular sets of genes as they develop, determining just what developmental processes occur, where, and when.
In many cases the same gene is found to control the same developmental process in many, if not all, animals. For example, a gene in mice called Pax6 encodes a transcription factor that initiates development of the eye. Mice without a functional copy of this gene do not make the transcription factor and are eyeless. When a gene was discovered in fruit flies that caused the flies to lack eyes, this gene was found to have essentially the same DNA sequence as the mouse gene—the same Pax6 master regulator gene was responsible for triggering eye development in both insects and vertebrates. Indeed, when Swiss biologist Walter Gehring inserted the mouse version of Pax6 into the fruitfly genome, a compound eye (the multifaceted kind that flies have) was formed on the leg of the fly! It seems that although insects and vertebrates diverged from a common ancestor more than 500 million years ago, they still control their development with genes so similar that the vertebrate gene seems to function quite normally in the insect genome.
Pax6 plays this same role of releasing eye development in many other animals. Even marine ribbon worms use it to initiate development of their eye spots. The Pax6 genes of all these animals have similar gene sequences, suggesting that Pax6 acquired its evolutionary role in eye development only a single time more than 500 million years ago, in the common ancestor of all animals that use Pax6 today.
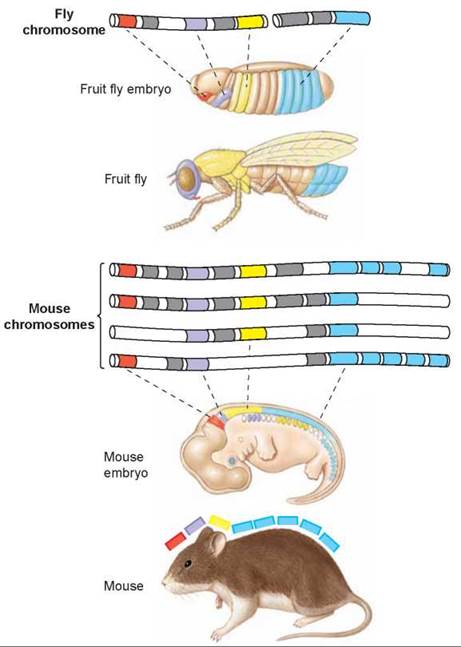
A more ancient master regulator gene called Hox determines basic body form. Hox genes appeared before the divergence of plants and animals; in plants they modulate shoot growth and leaf form, and in animals they establish the basic body plan.
All segmented animals appear to use organized clusters of Hox genes to control their development. After the sequential action of several "segmentation” genes, the body of these early embryos has a basically segmented body plan. This is true of the embryos of earthworms, fruit flies, mice— and human beings. The key to the further development of the animal body is to now give identity to each of the segments—to determine whether a particular segment will become back, or neck, or head, for example. In fruit flies and mice, similar clusters of Hox genes control this process. Flies have a single set of Hox genes, located on the same chromosome, while mice have four sets, each on a different chromosome (it appears the vertebrate genome underwent two entire duplications early in vertebrate evolution). In the illustration here, the genes are color-coded to match the parts of the body in which they are expressed.
How does a cluster of Hox genes work together to control segment development? Each Hox gene produces a protein with an identical 60-amino-acid segment that lets it bind to DNA as a transcription factor and, in so doing, activate the genes located where it binds. The differences between each Hox gene of a set determine where on DNA a Hox protein binds, and so which set of genes it activates.
Hox genes have also been found in clusters in radially symmetrical cnidarians such as hydra, suggesting that the ancestral Hox cluster preceded the divergence of radially and symmetrical animals in animal evolution.