MCAT Biology Review
Chapter 3: Embryogenesis and Development
Introduction
Is it a boy or a girl? It’s one of the most common questions asked of pregnant women. The suspense around learning the baby’s sex never fails to excite family and friends. For centuries, members of the elder generations have offered advice on how to predict the outcome or even plan it—Dangle a needle over your belly by a thread. Does it swing side to side or in circles?
In the modern world, needle dangling doesn’t carry the same credibility as technological advances like ultrasonography. This radiographic technique is performed by placing a probe that emits high-frequency sound waves near the tissue to be examined. The probe transduces an image onto a computer screen, which can be measured to determine gestational age, screen for multiple pregnancies or anomalies, and identify the baby’s sex. So how early can we determine a baby’s sex? All embryos begin female by default—that is, for a male fetus to develop, it must undergo not only masculinization via the gene product of SRY, but also defeminization. These processes occur (or don’t) around six to eight weeks postfertilization. Don’t expect any answers before 16 to 17 weeks, however, because ultrasonography equipment does not have high enough resolution yet to give immediate answers.
In this chapter, we’ll continue the discussion from the previous chapter by beginning with fertilization, the formation of a diploid zygote from the union of a sperm and an ovum. We’ll then follow development from this point until the birth of an autonomously breathing baby. We’ll examine how the cells of a developing human divide and differentiate. We’ll also explore some specific system differences that exist between developing fetuses and adults as we present an overview of the stages of pregnancy and childbirth.
3.1 Early Developmental Stages
In this first section, we explore development from the formation of a diploid zygote until neurulation, or the formation of the neural tube that will differentiate into the nervous system.
FERTILIZATION
As discussed in Chapter 2 of MCAT Biology Review, a secondary oocyte is ovulated from the follicle on approximately day 14 of the menstrual cycle. The secondary oocyte travels into the fallopian tube, where it can be fertilized up to 24 hours after ovulation. Fertilization, shown in Figure 3.1, usually occurs in the widest part of the fallopian tube, called the ampulla. When the sperm meets the secondary oocyte in the fallopian tube, it binds to the oocyte and releases acrosomal enzymes that enable the head of the sperm to penetrate through the corona radiata and zona pellucida. Once the first sperm comes into direct contact with the secondary oocyte’s cell membrane, it forms a tubelike structure known as the acrosomal apparatus, which extends to and penetrates the cell membrane. Its pronucleus may then freely enter the oocyte once meiosis II has come to completion. After penetration of the sperm through the cell membrane, the cortical reaction, a release of calcium ions, occurs. These calcium ions depolarize the membrane of the ovum, which serves two purposes: depolarization prevents fertilization of the ovum by multiple sperm cells, and the increased calcium concentration increases the metabolic rate of the newly formed diploid zygote. The now depolarized and impenetrable membrane is called the fertilization membrane.
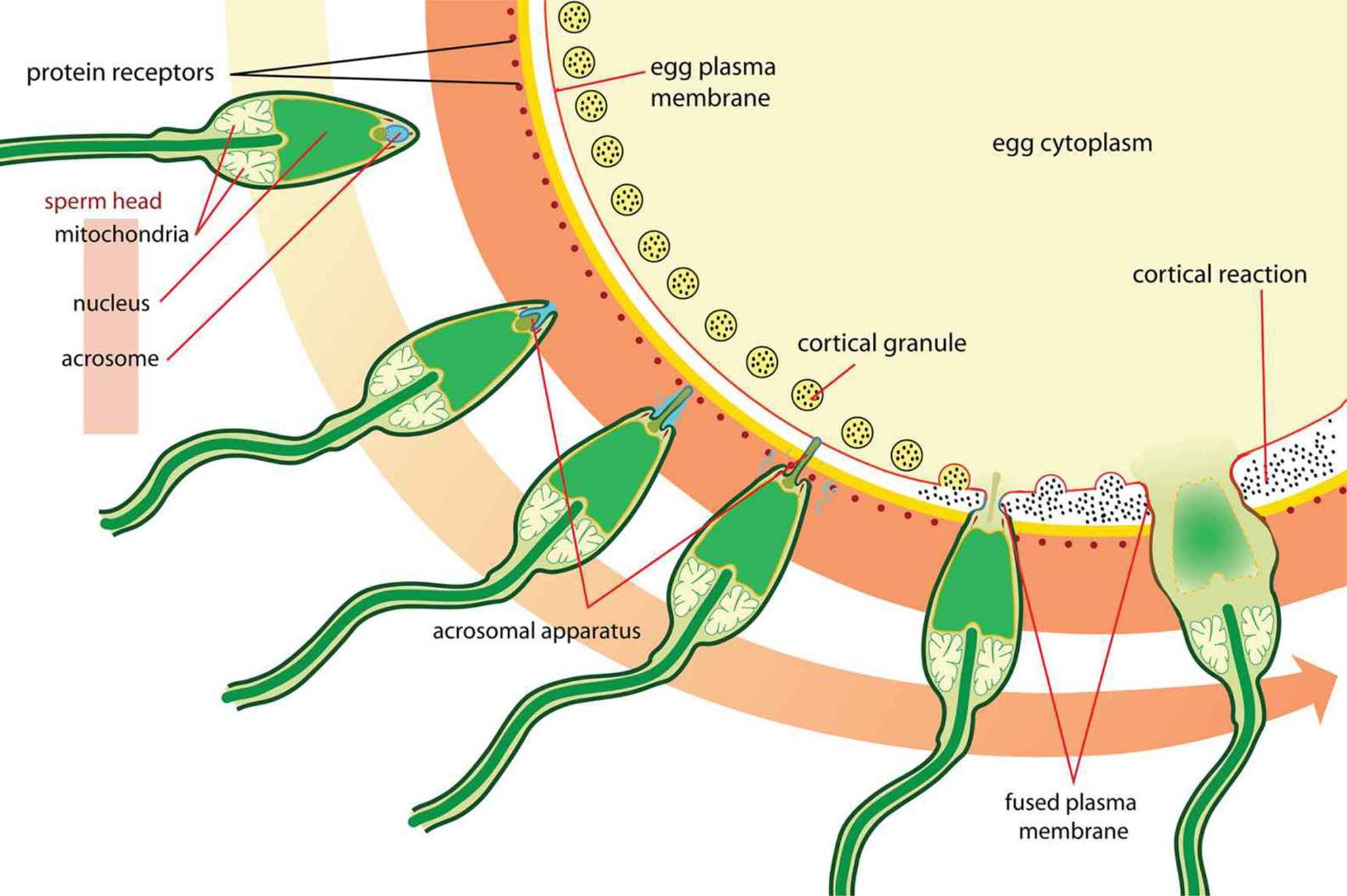 Figure 3.1. Fertilization
Figure 3.1. Fertilization
Twins
Twins can occur by two different mechanisms. Dizygotic, or fraternal twins, form from fertilization of two different eggs released during one ovulatory cycle by two different sperm. Each zygote will implant in the uterine wall, and each develops its own placenta, chorion, and amnion—these structures are discussed later in the chapter. If the zygotes implant close together, the placentas may fuse. Fraternal twins are no more genetically similar than any other pair of siblings.
Monozygotic, or identical twins, form when a single zygote splits into two. Because the genetic material is identical, so too will be the genomes of the offspring. If division is incomplete, conjoined twins may result, in which the two offspring are physically attached at some point. Monozygotic twins can be classified by the number of structures they share. Monochorionic/monoamniotic twins share the same amnion and chorion. Monochorionic/diamniotic twins each have their own amnion, but share the same chorion. Dichorionic/diamniotic twins each have their own amnions and chorions. Which type of twinning occurs is a result of when the separation occurred. As more gestational structures are shared, there are more risks as the fetuses grow and develop.
CLEAVAGE
After fertilization in the fallopian tubes, the zygote must travel to the uterus for implantation. If it arrives too late, there will no longer be an endometrium capable of supporting the embryo. In the process of moving to the uterus for implantation, the zygote undergoes rapid mitotic cell divisions in a process called cleavage. The first cleavage officially creates an embryo, as it nullifies one of the zygote’s defining characteristics: unicellularity. Although several rounds of mitosis occur, the total size of the embryo remains unchanged during the first few divisions, as shown in Figure 3.2. By dividing into progressively smaller cells, the cells increase two ratios: the nuclear-to-cytoplasmic (N:C) ratio and the surface area-to-volume ratio. Thus, the cells achieve increased area for gas and nutrient exchange relative to overall volume. There are two types of cleavage: indeterminate and determinate. Indeterminate cleavage results in cells that can still develop into complete organisms. In fact, monozygotic twins have identical genomes because they both originate from indeterminately cleaved cells of the same embryo. Determinate cleavage results in cells with fates that are, as the term implies, already determined. In other words, these cells are committed to differentiating into a certain type of cell.
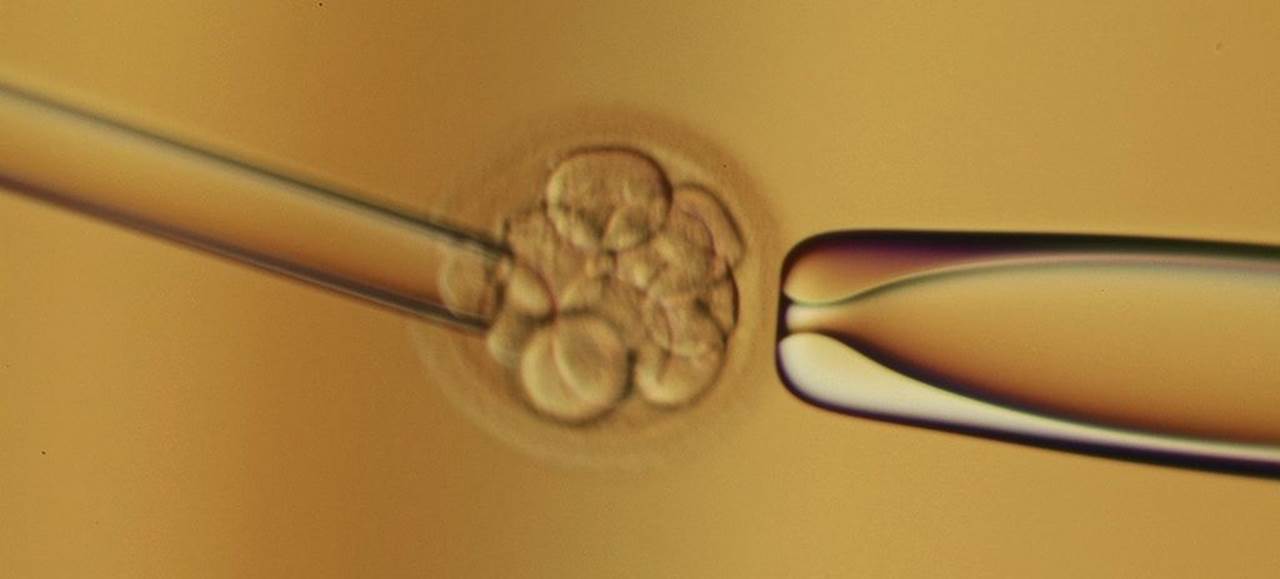 Figure 3.2. An 8-Cell Embryo The embryo has undergone three cleavage events at this point.
Figure 3.2. An 8-Cell Embryo The embryo has undergone three cleavage events at this point.
BLASTULATION
Several divisions later, the embryo becomes a solid mass of cells known as a morula, as shown in Figure 3.3. This term comes from the Latin word for mulberry, which might help us grasp what an embryo at this stage looks like.
 Figure 3.3. Morula The morula is a solid ball of cells.
Figure 3.3. Morula The morula is a solid ball of cells.
Once the morula is formed, it undergoes blastulation, which forms the blastula, a hollow ball of cells with a fluid-filled inner cavity known as a blastocoel. The mammalian blastula is known as a blastocyst and consists of two noteworthy cell groups, as shown in Figure 3.4: the trophoblast and inner cell mass. The trophoblast cells surround the blastocoel and give rise to the chorion and later the placenta, whereas the inner cell mass protrudes into the blastocoel and gives rise to the organism itself.
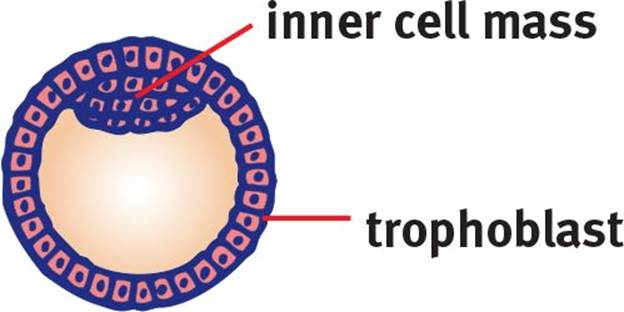 Figure 3.4. Blastula The blastula contains a fluid-filled cavity called the blastocoel.
Figure 3.4. Blastula The blastula contains a fluid-filled cavity called the blastocoel.
MNEMONIC
Remember that an embryo with a blasted-out cavity is a blastula.
Implantation
The blastula moves through the fallopian tube to the uterus, where it burrows into the endometrium. The trophoblast cells are specialized to create an interface between the maternal blood supply and the developing embryo. These trophoblastic cells give rise to the chorion, an extraembryonic membrane that develops into the placenta. The trophoblasts form chorionic villi, which are microscopic fingerlike projections that penetrate the endometrium. As these chorionic villi develop into the placenta, they support maternal–fetal gas exchange. The embryo is connected to the placenta by the umbilical cord, which consists of two arteries and one vein encased in a gelatinous substance. The vein carries freshly oxygenated blood rich with nutrients from the placenta to the embryo. The umbilical arteries carry deoxygenated blood and waste to the placenta for exchange.
REAL WORLD
Sometimes the blastula implants itself outside the uterus, a situation known as an ectopic pregnancy. Over 95% of ectopic pregnancies occur in the fallopian tube. Ectopic pregnancies are generally not viable because the narrow fallopian tube is not an environment in which an embryo can properly grow. If the embryo does not spontaneously abort, the tube may rupture, and a considerable amount of hemorrhaging may occur. In fact, a suspected ectopic pregnancy is often a surgical emergency.
Until the placenta is functional, the embryo is supported by the yolk sac. The yolk sac is also the site of early blood cell development. There are two other extraembryonic membranes that require discussion: the allantois and the amnion. The allantois is involved in early fluid exchange between the embryo and the yolk sac. Ultimately, the umbilical cord is formed from remnants of the yolk sac and the allantois. The allantois is surrounded by the amnion, which is a thin, tough membrane filled with amniotic fluid. This fluid serves as a shock absorber during pregnancy, lessening the impact of maternal motion on the developing embryo. In addition to forming the placenta, the chorion also forms an outer membrane around the amnion, adding an additional level of protection. The anatomy of these structures is shown in Figure 3.5.
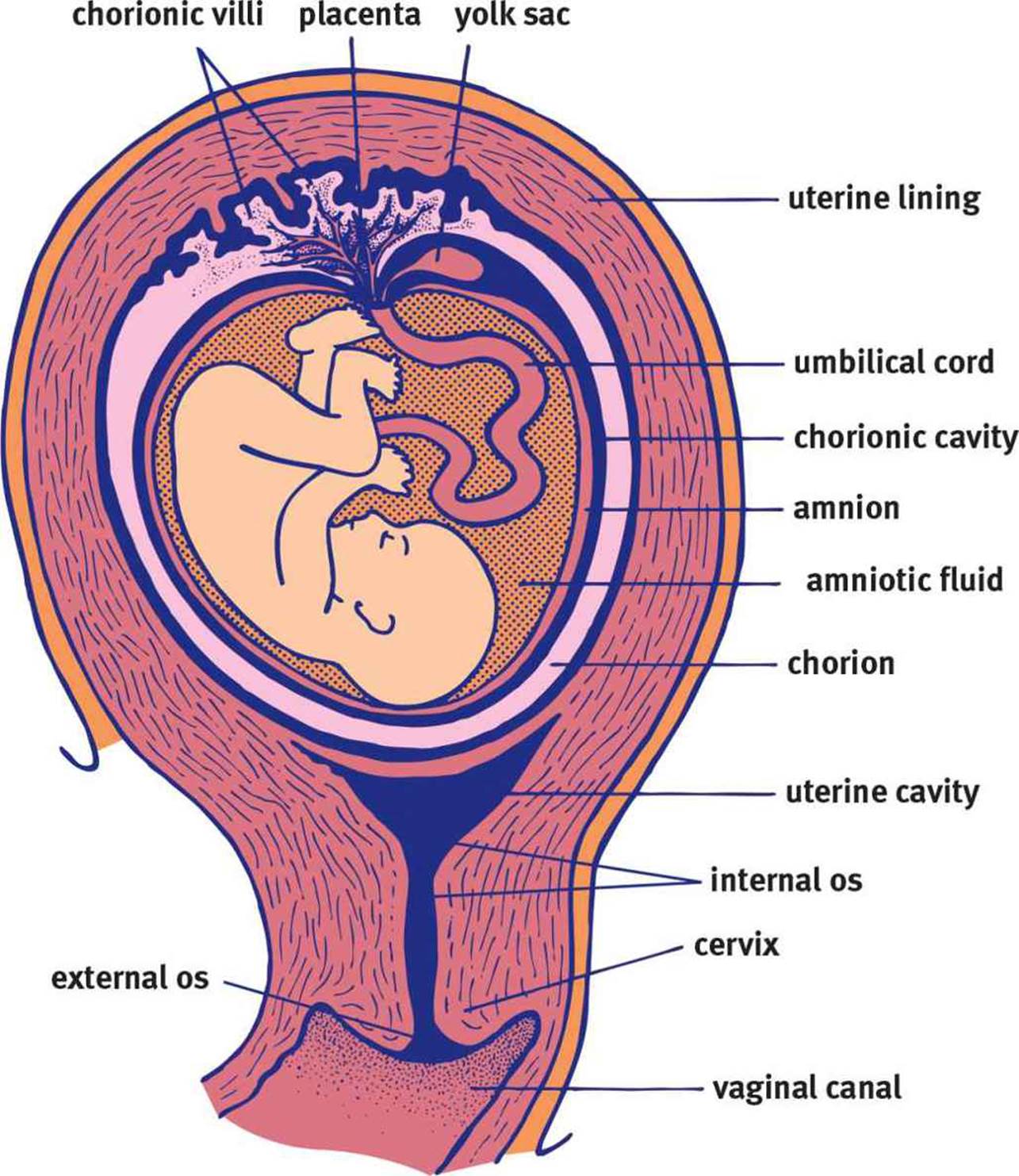 Figure 3.5. Anatomy of Pregnancy
Figure 3.5. Anatomy of Pregnancy
REAL WORLD
Amniocentesis is the process of aspirating amniotic fluid by inserting a thin needle into the amniotic sac. The amniotic fluid contains fetal cells that can be examined for chromosomal abnormalities as well as sex determination. Amniocentesis is recommended for pregnant women over 35 if earlier screening tests (blood tests and ultrasound) indicate a high chance of chromosomal abnormalities in the fetus. Women in this age group have a higher rate of meiotic nondisjunction, which can result in genetic aberrations such as Down syndrome.
GASTRULATION
Once the cell mass implants, it can begin further developmental processes such as gastrulation, the generation of three distinct cell layers. The early developmental processes up to this point are shown in Figure 3.6. Much of our knowledge of development comes from the study of other organisms, which have varying degrees of similarity to human development. In sea urchins, gastrulation begins with a small invagination in the blastula. Cells continue moving toward the invagination, resulting in elimination of the blastocoel. To visualize this, imagine inflating a balloon and then poking one of the sides with your finger. If you kept pushing, eventually the rubber from one side of the balloon would come into contact with the other side. If the two membranes could merge, as occurs in development, this would create a tube through the middle of the balloon. In living things, the result of this process is called a gastrula. The membrane invagination into the blastocoel is called the archenteron, which later develops into the gut. The opening of the archenteron is called the blastopore. In deuterostomes, such as humans, the blastopore develops into the anus. In protostomes, it develops into the mouth.
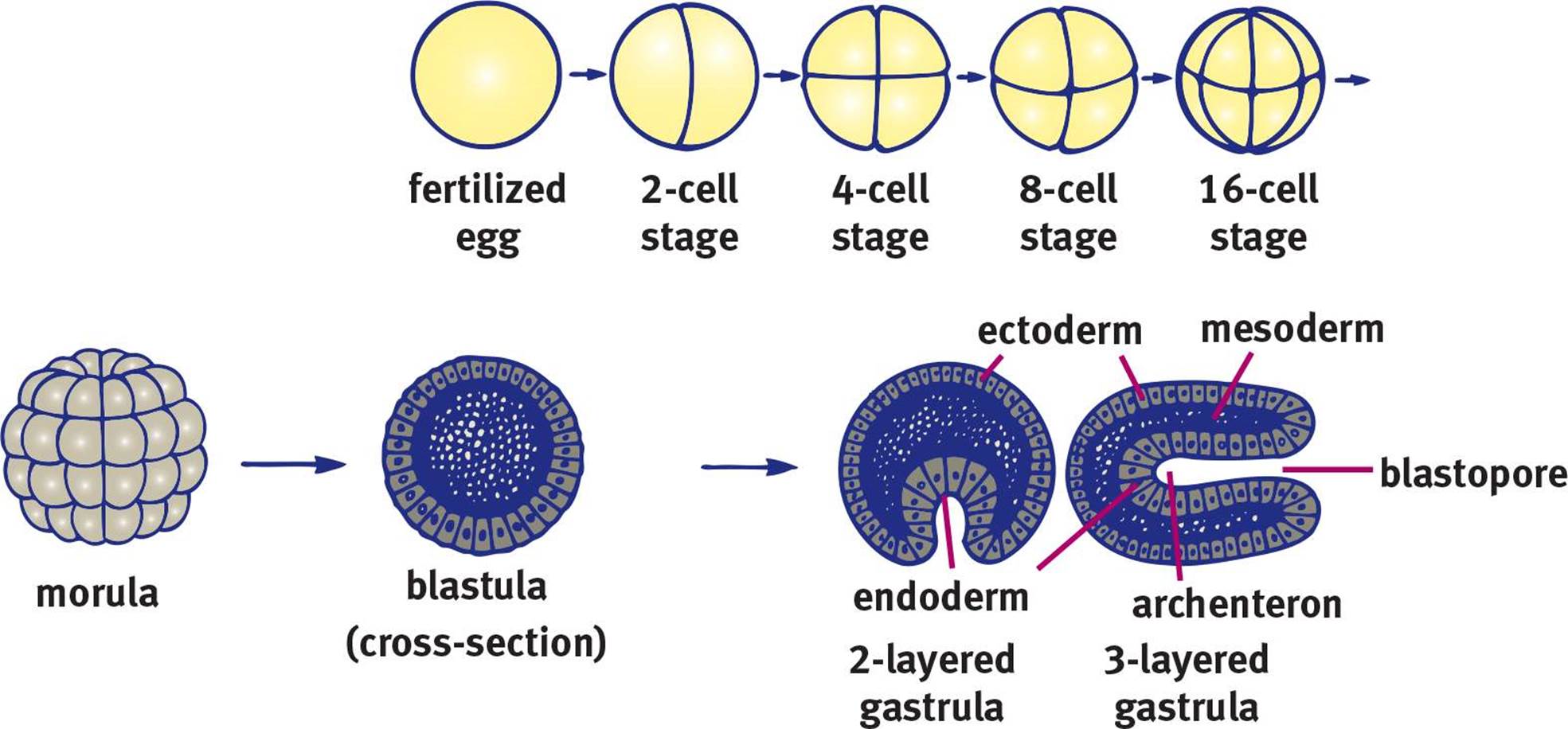 Figure 3.6. Early Stages of Embryonic Development
Figure 3.6. Early Stages of Embryonic Development
MNEMONIC
How can we remember the blastopore’s fate in protostomes vs. deuterostomes? Think about how parents talk to toddlers—deuterostome starts with deu, which looks like duo, meaning two. Thus, deuterostomes develop the anus—the orifice associated with “number two”—from the blastopore. Protostomes must start at the other end (the mouth).
Primary Germ Layers
Eventually, some cells will also migrate into what remains of the blastocoel. This establishes three layers of cells called primary germ layers.
The outermost layer is called the ectoderm and gives rise to the integument, including the epidermis, hair, nails, and the epithelia of the nose, mouth, and lower anal canal. The lens of the eye, nervous system (including adrenal medulla), and inner ear are also derived from ectoderm.
The middle layer is called the mesoderm and develops into several different systems including the musculoskeletal, circulatory, and most of the excretory systems. Mesoderm also gives rise to the gonads as well as the muscular and connective tissue layers of the digestive and respiratory systems and the adrenal cortex.
The innermost layer is called the endoderm and forms the epithelial linings of the digestive and respiratory tracts, including the lungs. The pancreas, thyroid, bladder, and distal urinary tracts, as well as parts of the liver, are derived from endoderm.
MNEMONIC
The primary germ layers:
· Ectoderm—“attracto”derm (things that attract us to others, such as cosmetic features and “smarts”)
· Mesoderm—“means”oderm (the means of getting around as an organism, such as bones and muscle; the means of getting around in the body, such as the circulatory system; the means of getting around, such as the gonads)
· Endoderm—linings of “endernal” organs (the digestive and respiratory tract, and accessory organs attached to these systems)
MCAT EXPERTISE
The MCAT likes to test on the dual embryonic origin of the adrenal glands. The adrenal cortex is derived from the mesoderm, but the adrenal medulla is derived from the ectoderm (because the adrenal medulla contains some nervous tissue).
Differentiation
So how is it that cells with the same genes are able to develop into such distinctly different cell types with highly specialized functions? Primarily, it is by selective transcription of the genome. In other words, only the genes needed for that particular cell type are transcribed. Thus, in pancreatic islet cells, the genes to produce specific hormones (insulin, glucagon, or somatostatin) are turned on, while these same genes are turned off in other cell types. Selective transcription is often related to the concept of induction, which is the ability of one group of cells to influence the fate of other nearby cells. This process is mediated by chemical substances called inducers, which diffuse from the organizing cells to the responsive cells. These chemicals are responsible for processes such as the guidance of neuronal axons. This process also ensures proximity of different cell types that work together within an organ.
NEURULATION
Once the three germ layers are formed, neurulation, or development of the nervous system, can begin. Remember that the nervous system is derived from the ectoderm. How, then, do cells originating on the surface of the embryo (ectoderm) end up inside the final organism? First, a rod of mesodermal cells known as the notochord forms along the long axis of the organism like a primitive spine (in fact, small remnants of notochord persist in the intervertebral discs between vertebrae). The notochord induces a group of overlying ectodermal cells to slide inward to form neural folds, which surround a neural groove, as shown in Figure 3.7. The neural folds grow toward one another until they fuse into a neural tube, which gives rise to the central nervous system. At the tip of each neural fold are neural crest cells. These cells migrate outward to form the peripheral nervous system (including the sensory ganglia, autonomic ganglia, adrenal medulla, and Schwann cells) as well as specific cell types in other tissues (such as calcitonin-producing cells of the thyroid, melanocytes in the skin, and others). Finally, ectodermal cells will migrate over the neural tube and crests to cover the rudimentary nervous system.
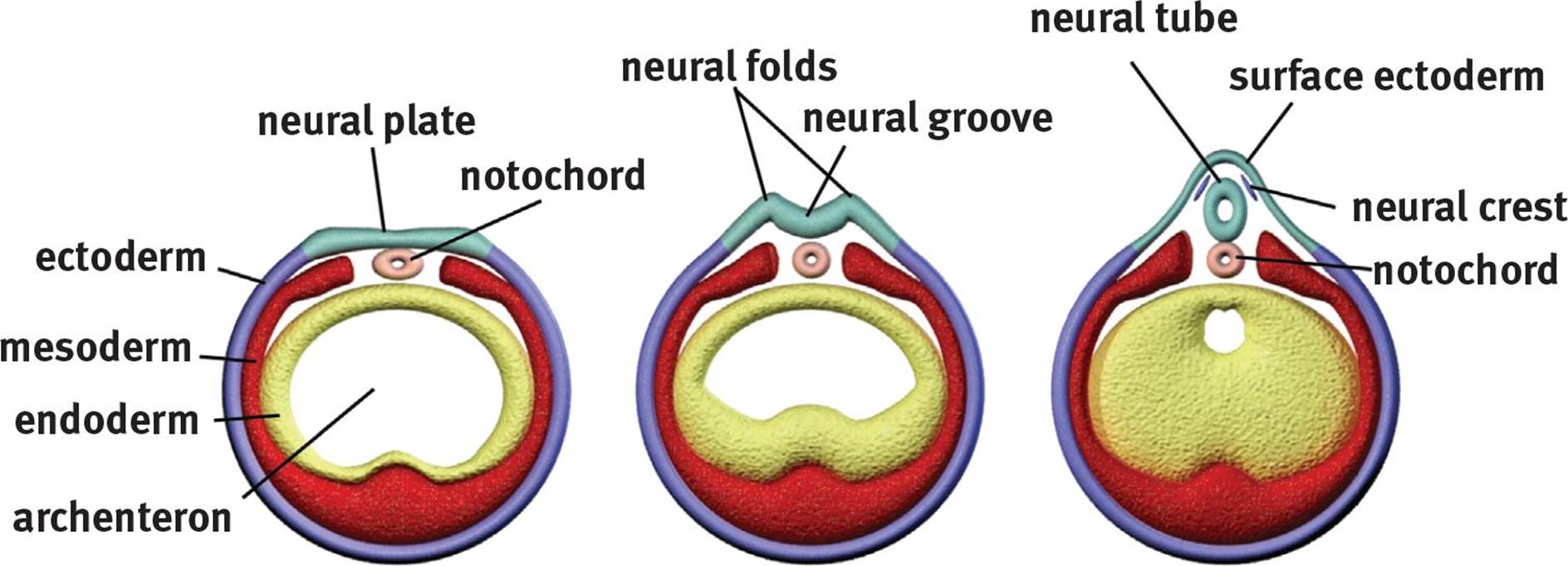 Figure 3.7. Formation of the Neural Tube
Figure 3.7. Formation of the Neural Tube
REAL WORLD
Failure of the neural tube to close results in either spina bifida (in which some or all of the spinal cord may be exposed to the outside world) or anencephaly (in which the brain fails to develop and the skull is left open). The severity of spina bifida’s effects range from no significant distress to death, whereas anencephaly is universally fatal. Women who wish to conceive are encouraged to take folate (folic acid) to prevent this complication; it is recommended that all women of childbearing age supplement their diets with folate in advance of getting pregnant because neurulation often occurs before pregnancy is detected.
PROBLEMS IN EARLY DEVELOPMENT
Early development is a highly sensitive time during the development of a human being. During this stage, as the germ layers are forming and then organogenesis (the production of organs) begins, teratogens may have far-reaching and highly detrimental effects. Teratogens are substances that interfere with development, causing defects or even death of the developing embryo. However, each teratogen will not have the same effect on every embryo or fetus. It is believed that the genetics of the individual embryo influences the effects of the teratogen. In addition to genetics, the route of exposure, length of exposure, rate of placental transmission of the teratogen, and the exact identity of the teratogen will also affect the overall outcome. Some common teratogens include alcohol, prescription drugs, viruses, bacteria, and environmental chemicals including polycyclic aromatic hydrocarbons.
In addition to teratogens, development can also be influenced by maternal health. Certain conditions may cause changes in the overall physiology of the mother, resulting in overexposure or underexposure of the embryo or fetus to certain chemicals. For example, diabetic mothers with hyperglycemia (high blood glucose) can have poor birth outcomes. Overexposure to sugar in utero can lead to a fetus that is too large to be delivered and who suffers from hypoglycemia soon after birth (due to synthesizing very high levels of insulin to compensate). Maternal folic acid deficiency may prevent complete closure of the neural tube, resulting in spina bifida or anencephaly, in which parts of the nervous system are exposed to the outside world or covered with a thin membrane. However, like teratogens, maternal health issues can have variable effects on the developing fetus. Spina bifida may be so severe as to result in profound disability, or may be completely asymptomatic and only detected by a tuft of hair overlying the area. Overall, trends and associations can certainly be found between various environmental conditions and genes during development; however, it is somewhat unpredictable and highly variable.
MCAT Concept Check 3.1:
Before you move on, assess your understanding of the material with these questions.
1. What is the difference between determinate and indeterminate cleavage?
· Determinate cleavage:
· Indeterminate cleavage:
2. From zygote to gastrula, what are the various stages of development?
3. During which stage of development does implantation occur?
4. What are the primary germ layers, and what organs are formed from each?
|
Germ Layer |
Organs |
5. What is induction?
6. What tissues do neural crest cells develop into?