5 Steps to a 5: AP Biology - Mark Anestis 2021
STEP 4 Review the Knowledge You Need to Score High
CHAPTER 9 Heredity
Exam Weight: 8—11%
IN THIS CHAPTER
Summary: This chapter examines Mendel’s fundamental laws (law of segregation, law of independent assortment, and law of dominance) as well as some classic exceptions to these laws (intermediate inheritance, multiple alleles, polygenic traits, epistasis, and pleiotropy). This chapter also covers linkage (sex linkage, gene linkage, and linkage maps), and chromosomal errors such as nondisjunction, deletions, duplications, translocations, and inversion.

Key Ideas
![]() Crossing over occurs during prophase I of meiosis.
Crossing over occurs during prophase I of meiosis.
![]() Sources of cell variation: crossover, 2n possible gametes, and random pairing of gametes.
Sources of cell variation: crossover, 2n possible gametes, and random pairing of gametes.
![]() Law of segregation: the two alleles for a trait separate during the formation of gametes—one to each gamete.
Law of segregation: the two alleles for a trait separate during the formation of gametes—one to each gamete.
![]() Law of independent assortment: inheritance of one trait does not interfere with the inheritance of another trait.
Law of independent assortment: inheritance of one trait does not interfere with the inheritance of another trait.
![]() Law of dominance: if two opposite pure-breeding varieties are crossed (BB × bb), all offspring resemble the BB parent.
Law of dominance: if two opposite pure-breeding varieties are crossed (BB × bb), all offspring resemble the BB parent.
![]() Linked genes that lie along the same chromosome do not follow the law of independent assortment.
Linked genes that lie along the same chromosome do not follow the law of independent assortment.
![]() Autosomal recessive disorders: Tay-Sachs, cystic fibrosis, sickle cell anemia, phenylketonuria.
Autosomal recessive disorders: Tay-Sachs, cystic fibrosis, sickle cell anemia, phenylketonuria.
![]() Autosomal dominant disorders: Huntington, achondroplasia.
Autosomal dominant disorders: Huntington, achondroplasia.
![]() Nondisjunction errors: Down, Klinefelter, Turner syndromes.
Nondisjunction errors: Down, Klinefelter, Turner syndromes.
Introduction
How many times have you heard someone say as they look at a baby, “Awwww, he looks like his daddy” or “She has her mother’s eyes”? What exactly is it that causes an infant to look like his or her parents? This question is the basis of the study of heredity—the study of the passing of traits from generation to generation. Its basic premise is that offspring are more like their parents than less closely related individuals.
In this chapter, we will begin by discussing meiosis. We will then discuss some terms that will prove important to your study of heredity. This is followed by an examination of Mendel’s law of segregation and the law of independent assortment, including how they were discovered and how they can be applied. We will examine the law of dominance, which arose from Mendel’s work, and we will also discuss some exceptions to Mendel’s fundamental laws such as intermediate inheritance (incomplete dominance and codominance), multiple alleles, polygenic traits, epistasis, and pleiotropy.
In the next section, we will examine Thomas Morgan’s work on fruit flies, which paved the way for the discovery of linked genes, genetic recombination, and sex-linked inheritance. This discussion concludes with a look at gene linkage and linkage maps.
Finally, since chromosomes carry the vital genes necessary for proper development and passage of hereditary material from one generation to the next, it is important to discuss the types of chromosomal errors that can occur during reproduction. This includes the various forms of nondisjunction, or the improper separation of chromosomes during meiosis (which leads to an abnormal number of chromosomes in offspring). The chapter concludes with an examination of the other major types of chromosomal errors: deletions, duplications, translocations, and inversions.
Haploid Versus Diploid Organisms
One thing that is often a major source of confusion for some of my students is the distinction between being haploid and being diploid. Let’s start with a definition of the terms:
A haploid (n) organism is one that has only one copy of each type of chromosome. In humans, this refers to a cell that has one copy of each type of homologous chromosome.
A diploid (2n) organism is one that has two copies of each type of chromosome. In humans, this refers to the pairs of homologous chromosomes.
During the discussion of meiosis below, the terms haploid and diploid will be used often. Whenever we say “2n,” or diploid, we are referring to an organism that contains two full sets of chromosomes. The letter n is used to represent the number of sets of chromosomes. So if an organism is said to have 4n chromosomes, this means that it has four complete sets of chromosomes. Humans are diploid, and consist of 2n chromosomes at all times except as gametes, when they are n. Humans have 23 different chromosomes; there are two full sets of these 23 chromosomes, one from each parent, for a total of 46 chromosomes. Human sex cells have 23 chromosomes each.
Meiosis
Now that we have armed you with the knowledge of the distinction between haploid and diploid, it is time to dive into the topic of meiosis, which occurs during the process of sexual reproduction. A cell destined to undergo meiosis goes through the cell cycle, synthesizing a second copy of DNA just like mitotic cells. But after G2, the cell instead enters meiosis, which consists of two cell divisions, not one. The second cell division exists because the gametes to be formed from meiosis must be haploid. This is because they are going to join with another haploid gamete at conception to produce the diploid zygote. Meiosis is like a two-part made-for-TV miniseries. It has two acts: meiosis I and meiosis II. Each of these two acts is divided into four steps, reminiscent of mitosis: prophase, metaphase, anaphase, and telophase.
SYI-1
Heritable information provides for continuity of life.
Homologous chromosomes resemble one another in shape, size, function, and the genetic information they contain. In humans, the 46 chromosomes are divided into 23 homologous pairs. One member of each pair comes from an individual’s mother, and the other member comes from the father. Meiosis I is the separation of the homologous pairs into two separate cells. Meiosis II is the separation of the duplicated sister chromatids into chromosomes. As a result, a single meiotic cycle produces four cells from a single cell. The cells produced during meiosis in the human life cycle are called gametes.
Again, the AP Biology exam is not going to test your mastery of the minute details of the meiotic process. However, a general understanding of the various steps is important:
Meiosis I
Prophase I. Each chromosome pairs with its homolog. Crossover (synapsis) occurs in this phase. The nuclear envelope breaks apart, and spindle apparatus begins to form.
Metaphase I. Chromosomes align along the metaphase plate matched with their homologous partner. This stage ends with the separation of the homologous pairs.
Anaphase I. Separated homologous pairs move to opposite poles of the cell.
Telophase I. Nuclear membrane reforms; the process of cytoplasmic division begins.
Cytokinesis. After the daughter cells split, the two newly formed cells are haploid (n).
As discussed earlier, meiosis consists of a single synthesis period during which the DNA is replicated, followed by two acts of cell division. With the completion of the first cell division, meiosis I, the cells are haploid because they no longer consist of two full sets of chromosomes. Each cell has one of the duplicated chromatid pairs from each homologous pair. The cell then enters meiosis II.
Meiosis II
Prophase II. The nuclear envelope breaks apart, and spindle apparatus begins to form.
Metaphase II. Sister chromatids line up along the equator of the cell.
Anaphase II. Sister chromatids split apart and are called chromosomes as they are pulled to the poles.
Telophase II. The nuclei and the nucleoli for the newly split cells return.
Cytokinesis. Newly formed daughter cells physically divide (See Figure 9.1).
In humans, the process of gamete formation is different in women and men. In men, spermatogenesis leads to the production of four haploid sperm during each meiotic cycle. In women, the process is called oogenesis. It is a trickier process than spermatogenesis, and each complete meiotic cycle leads to the production of a single ovum, or egg. After meiosis I in females, one cell receives half the genetic information and the majority of the cytoplasm of the parent cell. The other cell, the polar body, simply receives half of the genetic information and is cast away. During meiosis II, the remaining cell divides a second time, and forms a polar body that is cast away, and a single haploid ovum that contains half the genetic information and nearly all the cytoplasm of the original parent cell. The excess cytoplasm is required for proper growth of the embryo after fertilization. Thus, the process of oogenesis produces two polar bodies and a single haploid ovum.
To review, why is it important to produce haploid gametes during meiosis? During fertilization, a sperm (n) will meet up with an egg (n), to produce a diploid zygote (2n). If either the sperm or the egg were diploid, then the offspring produced during sexual reproduction would contain more chromosomes than the parent organism. Meiosis circumvents this problem by producing gametes that are haploid and consist of one copy of each type of chromosome. During fertilization between two gametes, each copy will match up with another copy of each type of chromosome to form the diploid zygote.
Before moving on, there are a few important distinctions between meiosis and mitosis that should be emphasized. In meiosis during prophase I, the homologous pairs join together. This matching of chromosomes into homologous pairs does not occur in mitosis. In mitosis, the 46 chromosomes simply align along the metaphase plate alone.
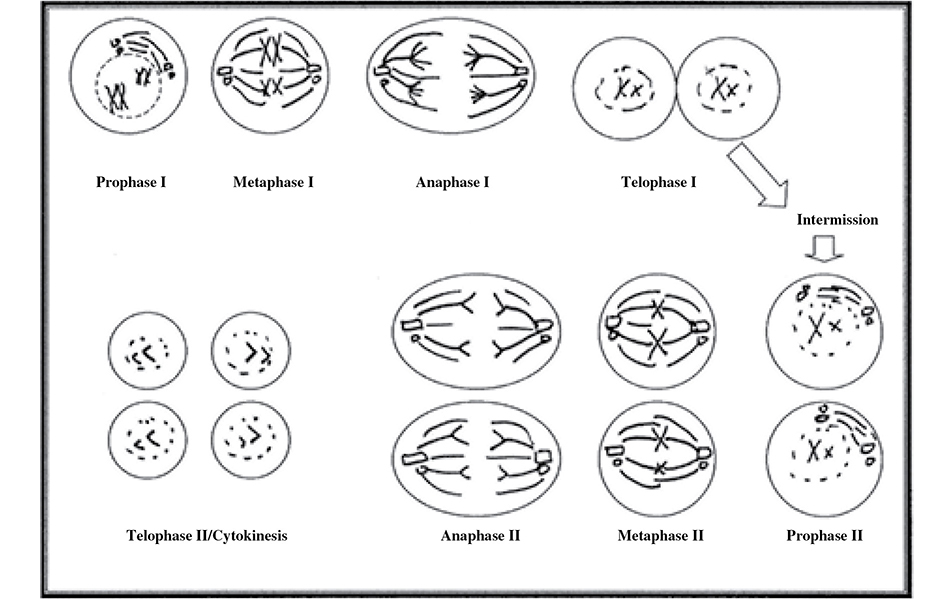
Figure 9.1 The stages of meiosis. Pictorial representation of the stages of meiosis I and II.

An event of major importance that occurs during meiosis that does not occur during mitosis is known as crossover (also known as crossing over) (Figure 9.2. When the homologous pairs match up during prophase I of meiosis, complementary pieces from the two homologous chromosomes wrap around each other and are exchanged between the chromosomes. Imagine that chromosome A is the homologous partner for chromosome B. When they pair up during prophase I, a piece of chromosome A containing a certain stretch of genes can be exchanged for the piece of chromosome B containing the same genetic information. This is one of the mechanisms that allows offspring to differ from their parents. Remember that crossing over occurs between the homologous chromosome pairs, not the sister chromatids.
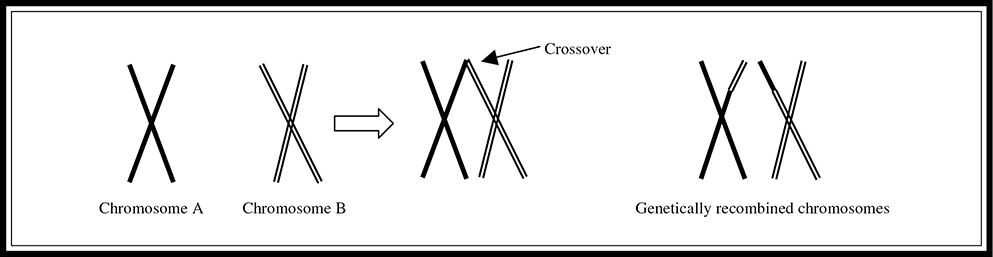
Figure 9.2 Crossover.
Some Important Terms to Know
The following is a list of terms that will help in your understanding of heredity:
Allele: a variant of a gene for a particular character. For example, two alleles for fur color could be B (dominant) and b (recessive).
F1: the first generation of offspring, or the first “filial” generation in a genetic cross.
F2: the second generation of offspring, or the second “filial” generation in a genetic cross.
Genotype: an organism’s genetic makeup for a given trait. A simple example of this could involve fur color where B represents the allele for brown and b represents the allele for black. The possible genotypes include homozygous brown (BB), heterozygous brown (Bb), and homozygous black (bb).
Heterozygous (hybrid): an individual is heterozygous (or a hybrid) for a gene if the two alleles are different (Bb).
Homozygous (pure): an individual is homozygous for a gene if both of the given alleles are the same (BB or bb).
Karyotype: a chart that organizes chromosomes in relation to number, size, and type.
Nondisjunction: the improper separation of chromosomes during meiosis, which leads to an abnormal number of chromosomes in offspring. A few classic examples of non-disjunction-related syndromes are Down, Turner, and Klinefelter syndromes.
P1: the parent generation in a genetic cross.
Phenotype: the physical expression of the trait associated with a particular genotype. Some examples of the phenotypes for Mendel’s peas were round or wrinkled, green or yellow, purple flower or white flower.
Sources of Cell Variation
What makes us different from our parents? Why do some people look amazingly like their parents while others do not? The process of cell division provides ample opportunity for variation. Remember that during meiosis, homologous chromosome pairs align together along the metaphase plate. This alignment is a completely random process, and there is a 50 percent chance that the chromosome in the pair from the individual’s mother will go to one side, and a 50 percent chance that the chromosome in the pair from the individual’s father will go to that side. This is true for all the homologous pairs in an organism. This means that 2n possible gametes can form from any given set of n chromosomes. For example, in a 3-chromosome organism, there are 23 = 8 possible gametes. In humans, there are 23 homologous pairs. This comes out to 223 (8,388,608) different ways the gametes can separate during gametogenesis.
NYC teacher: “Knowing the sources of variation is important.”
Another source of variation during sexual reproduction is the random determination of which sperm meets up with which ovum. In humans, the sperm represents one of 223 possibilities from the male gamete factory; the ovum, one of 223 possibilities from the female gamete factory. All these factors combine to explain why siblings may look nothing like each other.
A third major source of variation during gamete formation is the crossover (or crossing over) that occurs during prophase I of meiosis. It is very important for you to remember that this process happens only during that stage of cell division. It does not occur in mitosis.
Mendel and His Peas
The person whose name is most often associated with heredity is Gregor Mendel. Mendel spent many years working with peas. It was a very strange hobby, indeed, but it proved quite useful to the world of science. He mated peas to produce offspring and recorded the phenotype results in order to determine how certain characters are inherited. A character is a genetically inherited characteristic that differs from person to person.
EVO-2
Organisms are linked by lines of descent from common ancestry.
Before he began his work in the 1850s, the accepted theory of inheritance was the “blending” hypothesis, which stated that the genes contributed by two parents mix as colors do. For example, a blue flower mixed with a yellow flower would produce a green flower. The exact genetic makeup of each parent could never be recovered; the genes would be as inseparable as the blended colors. Mendel used plant experiments to test this hypothesis and developed his two fundamental theories: the law of segregation and the law of independent assortment.
SYI-1
Heritable information provides for continuity of life.
When Mendel was observing a single character during a mating, he was doing something called a monohybrid cross—a cross that involves a single character in which both parents are heterozygous (Bb × Bb). A monohybrid cross between heterozygous gametes gives a 3 : 1 phenotype ratio in the offspring (Figure 9.3. As you can see in Figure 9.3, an offspring is three times more likely to express the dominant B trait than the recessive b trait.
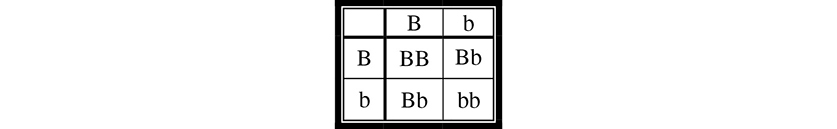
Figure 9.3 Monohybrid cross.
Mendel also experimented with multiple characters simultaneously. The crossing of two different hybrid characters is termed a dihybrid cross (BbRr × BbRr). A dihybrid cross between heterozygous gametes gives a 9 : 3 : 3 : 1 phenotype ratio in the offspring (Figure 9.4.
Figure 9.4 Dihybrid cross.
From his experiments, Mendel developed two major hereditary laws: the law of segregation and the law of independent assortment.
The law of segregation. Every organism carries pairs of factors, called alleles, for each trait, and the members of the pair segregate (separate) during the formation of gametes. For example, if an individual is Bb for eye color, during gamete formation, one gamete would receive a B, and the other made from that cell would receive a b.
The law of independent assortment. Members of each pair of factors are distributed independently when the gametes are formed. Quite simply, inheritance of one trait or characteristic does not interfere with inheritance of another trait. For example, if an individual is BbRr for two genes, gametes formed during meiosis could contain BR, Br, bR, or br. The B and b alleles assort independently of the R and r alleles.
The law of dominance. Also based on Mendel’s work, this states that when two opposite pure-breeding varieties (homozygous dominant vs. homozygous recessive) of an organism are crossed, all the offspring resemble one parent. This is referred to as the dominant trait. The variety that is hidden is referred to as the recessive trait.
It is time for you to answer a question for us (of course, we have no way of knowing whether or how you will answer this question): Can the phenotype of an organism be determined from simple observation? Yes—just look at the organism and determine whether it is tall or short, has blue eyes or brown eyes, and so on. However, the genotype of an organism cannot always be determined from simple observation. In the case of a recessive trait, the genotype is known. If a person has blue eyes (recessive to brown), the genotype is bb. But if that person has brown eyes, you cannot be sure if the genotype is Bb or BB—the individual can be either homozygous dominant or heterozygous dominant. To determine the exact genotype, you must run an experiment called a test cross. Geneticists breed the organism whose genotype is unknown with an organism that is homozygous recessive for the trait. This results in offspring with observable phenotypes. If the unknown genotype is heterozygous, probability indicates one-half of the offspring should express the recessive phenotype. If the unknown genotype is homo-zygous dominant, all the organism’s offspring should express the dominant trait. Of course, such experiments are not done on humans.
Remember me!
Mendel discovered many statistical laws of heredity. He learned that a monohybrid cross such as Yy × Yy will result in a phenotype ratio of 3:1 in favor of the dominant trait. He learned that a dihybrid cross, such as YyRr × YyRr, will result in a phenotype ratio of 9:3:3:1 (9 RY, 3 rY, 3 Ry, 1 ry). These two ratios, when they appear in genetic analysis problems, imply mono- and dihybrid crosses.
Non-Mendelian Genetics
Gregor Mendel’s work with pea plants and their genes was not able to account for many patterns of inheritance that occur in life (sex-linked traits, incomplete dominance, and codominance, to name a few). The observed phenotypes of these traits differ from the predicted ratios.
SYI-3
Naturally occurring diversity among and between components within biological systems affects interactions with the environment.
Non-nuclear inheritance, in which offspring get DNA only from the male or female parent, does not follow the Mendelian pattern of inheritance. For example, in humans, children get mitochondrial DNA from only the mother and not the father.
Intermediate Inheritance
The inheritance of traits is not always as simple as Mendel’s pea experiments seem to indicate. Traits are not always dominant or recessive, and phenotype ratios are not always 9:3:3:1 or 3:1. Mendel’s experiments did not account for something called intermediate inheritance, in which an individual heterozygous for a trait (Yy) shows characteristics not exactly like either parent. The phenotype is a “mixture” of both of the parents’ genetic input. There are two major types of intermediate inheritance:
Marcy (college freshman): “Understanding this concept is worth 2 points on the exam.”
1. Incomplete dominance or “blending inheritance”
2. Codominance
Incomplete Dominance (“Blending Inheritance”)
In incomplete dominance (“blending inheritance”), the heterozygous genotype produces an “intermediate” phenotype rather than the dominant phenotype; neither allele dominates the other. A classic example of incomplete dominance is flower color in snapdragons—crossing a snapdragon plant that has red flowers with one that has white flowers yields offspring with pink flowers.
One genetic condition in humans that exhibits incomplete dominance is hyper-cholesterolemia—a recessive disorder (hh) that causes cholesterol levels to be many times higher than normal and can lead to heart attacks in children as young as 2 years old. Those who are HH tend to have normal cholesterol levels, and those who are Hh have cholesterol levels somewhere in between the two extremes. As with many conditions, the environment plays a major role in how genetic conditions express themselves. Thus, people who are HH do not necessarily have normal cholesterol levels if, for example, they have poor diet or exercise habits.
One important side note—try not to confuse the terms blending “hypothesis” and blending “inheritance.” The latter is another name for incomplete dominance, whereas the former was the theory on heredity before Mendel worked his magic. The blending “hypothesis” says that the HH and hh extremes can never be retrieved. In reality, and according to blending inheritance, if you were to cross two Hh individuals, the offspring could still be HH or hh, which the blending “hypothesis” says cannot happen once the blending has occurred.
Codominance
Codominance is the situation in which both alleles express themselves fully in a heterozygous organism. A good example of codominance involves the human blood groups: M, N, and MN. Individuals with group M blood have the M glycoprotein on the surface of the blood cell; individuals with group N blood have N glycoproteins on the blood cell; and those with group MN blood have both. This is not incomplete dominance because both alleles are fully expressed in the phenotype—they are codominant.
Other Forms of Inheritance
Polygenic Traits
Another interesting form of inheritance involves polygenic traits, or traits that are affected by more than one gene. Eye color is an example of a polygenic trait. The tone (color), amount (blue eyes have less than brown eyes), and position (how evenly distributed the pigment is) of pigment all play a role in determining eye color. Each of these characteristics is determined by separate genes. Another example of this phenomenon is skin color, which is determined by at least three different genes working together to produce a wide range of possible skin tones.
Multiple Alleles
Many monogenic traits (traits expressed via a single gene) correspond to two alleles, one dominant and one recessive. Other traits, however, involve more than two alleles. A classic example of such a trait is the human blood type. On the most simplistic level, there are four major blood types: A, B, AB, and O. They are named based on the presence or absence of certain antigens on the surface of the red blood cells. The gene for blood type has three possible alleles (multiple alleles): IA, which causes antigens A to be produced on the surface of the red blood cell; IB, which causes antigens B to be produced; and i, which causes no antigens to be produced. The following are the possible genotypes for human blood type: IAi (type A), IAIA (type A), IBi (type B), IBIB (type B), IAIB (type AB), ii (type O). Type AB blood displays the codominance of blood type. As we saw in MN blood groups, both the A and the B alleles succeed in their mission—their antigens appear on the surface of the red blood cell (Figure 9.5. Analyzing blood type can be really complex becausehuman blood types involve not only multiple alleles (IA, IB, and i) and codominance (type AB blood), but classic dominance of IA and IB over i as well.
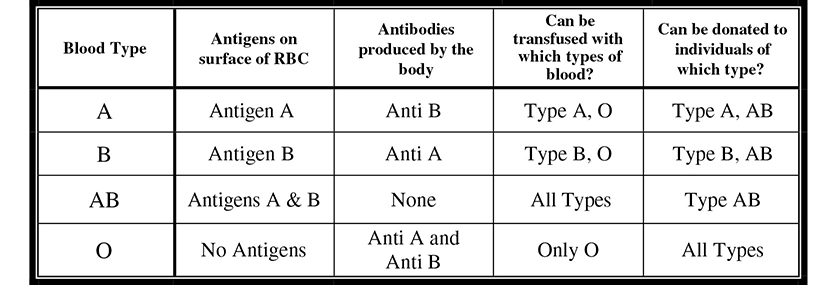
Figure 9.5 Several human blood type characteristics.
If you have ever watched an episode of ER on television, you have heard one of the doctors frantically scream, “We need to type her and bring some O blood down here stat!” Why is it important for the physician to determine what type of blood the patient has, and why is it okay to give the patient O blood in the meantime? People with type A blood produce anti-B antibodies because the B antigen that is present on type B and type AB blood is a foreign molecule to someone with type A blood. This is simply the body’s defense mechanism doing its job. Following the same logic, those with type B blood make anti-A antibodies, and those with type O blood make anti-A and anti-B antibodies. People who are type AB make none, and are therefore the universal acceptor of blood. It is important to find out what kind of blood a person has because if you give type B blood to a person with type A blood, the recipient will have an immune response to the transfused blood. Why is O blood given while they wait to see what blood type the patient is? This is because type O blood has neither antigen on the surface of red blood cells. People with type O blood are universal donors because few people will have an adverse reaction to type O blood.
Epistasis
In epistasis the expression of one gene affects the expression of another gene. A classic example of epistasis involves the coat color of mice. Black is dominant over brown, and brown fur has the genotype bb. There is also another gene locus independent of the coat color gene that controls the deposition of pigment in the fur. If a mouse has a dominant allele of this pigment gene (Cc or CC), it leads to pigment deposition and the coloring of the fur according to the coat color gene’s instructions. If a mouse is double recessive for this trait (cc), it will have white fur no matter what the coat color gene wants because it will not put any pigment into the fur. It is almost as if the pigment gene were overruling the coat color gene. If you mate two black mice that are BbCc, the ratio of phenotypes in the offspring would not be the 9 : 3 : 3 : 1 ratio that Mendel predicts, but rather 9 : 4 : 3 black : white : brown because the epistatic gene alters the phenotype.
Pleiotropy
In pleiotropy, a single gene has multiple effects on an organism. A good example of pleiotropy is the mutation that causes sickle cell anemia. This single gene mutation “sickles” the blood cells, leading to systemic symptoms such as heart, lung, and kidney damage; muscle pain; weakness; and generalized fatigue. The problems do not stop there; these symptoms can lead to disastrous side effects such as kidney failure. The mutation of a single gene wreaks havoc on the system as a whole.
Sex Determination and Sex Linkage
Mendel was not the only one to make progress in the field of heredity. In the early 1900s, Thomas Morgan made key discoveries regarding sex linkage and linked genes.
In human cells, all chromosomes occur in structurally identical pairs except for two very important ones: the sex chromosomes, X and Y. Women have two structurally identical X chromosomes. Men have one X and one Y.
Sex-Linked Traits
Morgan experimented with a quick-breeding fruit fly species. The fruit flies had four pairs of chromosomes: three autosomal pairs and one sex chromosome pair. An autosomal chromosome is one that is not directly involved in determining gender. In fruit flies, the more common phenotype for a trait is called the wild-type phenotype (e.g., red eyes). Traits that are different from the normal are called mutant phenotypes (e.g., white eyes). Morgan crossed a white-eyed male with a red-eyed female, and all the F1 offspring had red eyes. When he bred the F1 together, he obtained Mendel’s 3:1 ratio. But, there was a slight difference from what Mendel’s theories would predict—the white trait was restricted to the males. Morgan’s conclusion was that the gene for eye color is on the X chromosome. This means that the poor male flies get only a single copy, and if it is abnormal, they are abnormal. But, the lucky ladies have two copies and are normal even if one copy is not.
SYI-1
Heritable information provides for continuity of life.
It is this male—female sex chromosomes difference that allows for sex-linked conditions. If a gene for a recessive disease is present on the X chromosome, then a female must have two defective versions of the gene to show the disease while a male needs only one. This is so because males have no corresponding gene on the Y chromosome to help counter the negative effect of a recessive allele on the X chromosome. Thus, more males than females show recessive X-linked phenotypes. In a pedigree (see Figure 9.8 later in this chapter), a pattern of sex-linked disease will show the sons of carrier mothers with the disease.
The father plays no part in the passage of an X-linked gene to the male children of a couple. Fathers pass X-linked alleles to their daughters, but not to their sons. Do you understand why this is so? The father does not give an X chromosome to the male offspring because he is the one who provides the Y chromosome that makes his son a male. A mother can pass a sex-linked allele to both her daughters and sons because she can pass only X chromosomes to her offspring.
Emily (12th grader): “Be able to categorize diseases for this exam!”
Three common sex-linked disorders are Duchenne’s muscular dystrophy, hemophilia, and red-green colorblindness. Duchenne’s muscular dystrophy is a sex-linked disorder that is caused by the absence of an essential muscle protein. Its symptoms include a progressive loss of muscle strength and coordination. Hemophilia is caused by the absence of a protein vital to the clotting process. Individuals with this condition have difficulty clotting blood after even the smallest of wounds. Those most severely affected by the disease can bleed to death after the tiniest of injuries. Females with this condition rarely survive. People afflicted with red-green colorblindness are unable to distinguish between red and green colors. This condition is found primarily in males.
X Inactivation
Here is an important question for you to ponder while preparing for this exam: “Are all the cells in a female identical?”
The answer to this question is “No.” Females undergo a process called X inactivation. During the development of a female embryo, one of the two X chromosomes in each cell remains coiled as a Barr body whose genes are not expressed. A cell expresses the alleles only of the active X chromosome. X inactivation occurs separately in each cell and involves random inactivation of one of a female’s X chromosomes. But not all cells inactivate the same X. As a result, different cells will have different active X chromosomes.
Why don’t females always express X-linked diseases when this X inactivation occurs? Sometimes they do, but usually they have enough cells with a “good” copy of the allele to compensate for the presence of the recessive allele.
One last sex-related inheritance pattern that needs to be mentioned is holandric traits, which are traits inherited via the Y chromosome. An example of a holandric trait in humans is ear hair distribution.
Linkage and Gene Mapping
Each chromosome has hundreds of genes that tend to be inherited together because the chromosome is passed along as a unit. These are called linked genes. Linked genes lie on the same chromosome and do not follow Mendel’s law of independent assortment.
Morgan performed an experiment in which he looked at body color and wing size on his beloved fruit flies. The dominant alleles were G (gray) and V (normal wings); the recessive alleles were g (black) and v (vestigial wings). GgVv females were crossed with ggvv males. Mendel’s law of independent assortment predicts offspring of four different phenotypes in a 1:1:1:1 ratio. But that is not what Morgan found. Because the genes are linked, the gray/normal flies produce only GV or gv gametes. Thus, Morgan expected the ratio of offspring to be 1:1, half GgVv and half ggvv. Morgan found that there were more wild-type and double-mutant flies than independent assortment would predict, but surprisingly, some Gv and gV were also produced.
How did those other combinations result from the cross if the genes are linked? Crossover (also known as crossing over), a form of genetic recombination that occurs during prophase I of meiosis, led to their production. The less often this recombination occurs, the closer the genes must be on the chromosome. The farther apart two genes are on a chromosome, the more often crossover will occur. Recombination frequency can be used to determine how close two genes are on a chromosome through the creation of linkage maps, which we will look at next.
Linkage Maps
A linkage map is a genetic map put together using crossover frequencies. Another unit of measurement, the map unit (also known as centigram), is used to geographically relate the genes on the basis of these frequencies. One map unit is equal to a 1 percent crossover frequency. A linkage map does not provide the exact location of genes; it gives only the relative location. Imagine that you want to determine the relative locations of four genes: A, B, C, and D. You know that A crosses over with C 20 percent of the time, B crosses over with C 15 percent of the time, A crosses over with D 10 percent of the time, and D crosses over with B 5 percent of the time. From this information you can determine the sequence (Figure 9.6. Gene A must be 20 units from gene C. Gene B must be 15 units from C, but B could be 5 or 35 units from A. But, because you also know that A is 10 units from D and that D is 5 units from B, you can determine that B must be 5 units from A as well, if A is also to be 10 units from D. This gives you the sequence of genes as ABDC.
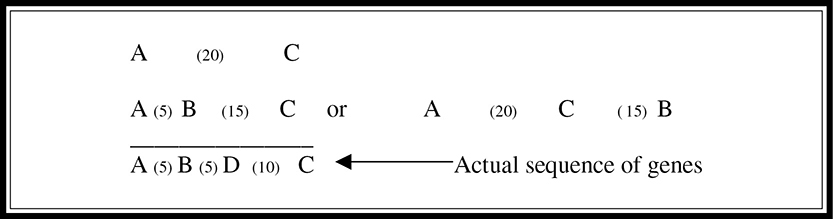
Figure 9.6 A genetic linkage map.
Heads or Tails?
Probability is a concept important to a full understanding of heredity and inheritance. What is the probability that a flipped coin will come up heads? You can answer that easily: ½. What is the probability that two coins flipped simultaneously will both be heads? This is a little harder—it is ½ × ½ = ¼. Take a look at Figure 9.7. The first time you toss the coin, there is a probability of ½ that it will land heads and ½ that it will land tails. When you toss it again, it again has a probability of ½ that it will land heads, and ½ that it will land tails. So in the figure, just concentrate on the ½ of the tosses that land heads. Of those, ½ of them will land heads the second time—or ½ of ½. Multiplied together, this results in the ¼ chance of getting heads twice with two coin tosses. This example illustrates the law of multiplication with probabilities. This law states that to determine the probability that two random events will occur in succession, you simply multiply the probability of the first event by the probability of the second event.
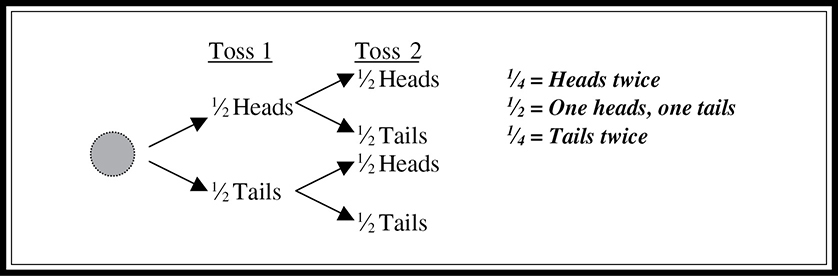
Figure 9.7 Probability in the law of multiplication.
This is the same thought process that we follow to understand Mendel’s law of segregation. If you are Aa for a trait, what is your chance of passing on the A? That’s right—½. If you are AaBb, what is the chance you pass on both A and B? Clever you are—you multiply ½ × ½ to get ¼.
Pedigrees
Pedigrees are family trees used to describe the genetic relationships within a family. Comprehension of the probability concept is important for a full understanding of pedigree analysis. Squares represent males, and circles are used for females. A horizontal line from male to female represents mates that have produced offspring. The offspring are listed below their parents from oldest to youngest. A fully shaded individual possesses the trait being studied. If the condition being studied is a monogenic recessive condition (rr), then those shaded gray have the genotype rr. If the condition being studied is a dominant condition (Rr or RR), then those that are unshaded have the genotype rr. A line through a symbol indicates that the person is deceased. A sample pedigree is shown in Figure 9.8.
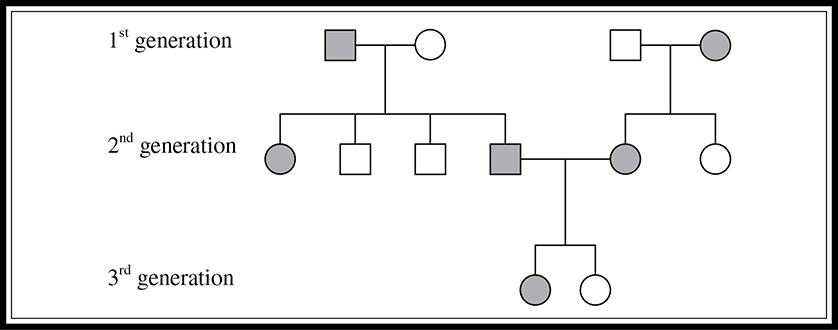
Figure 9.8 Schematic of a pedigree.
CT teacher: “Test almost always has two to three questions about this topic.”
Pedigrees can be used in many ways. One use is to determine the risk of parents passing certain conditions to their offspring. Imagine that two people want to have a child, and they both have a family history of a certain autosomal recessive condition (dd). Neither has the particular condition, but the man has a brother who died of the disease and the woman’s mother died of the disease at an older age. They want to know the probability of having a child with the condition. You must first determine the probability that each parent is a carrier, and then determine the probability of the parents having a child with the disease, given that they are carriers. See the pedigree in Figure 9.9.
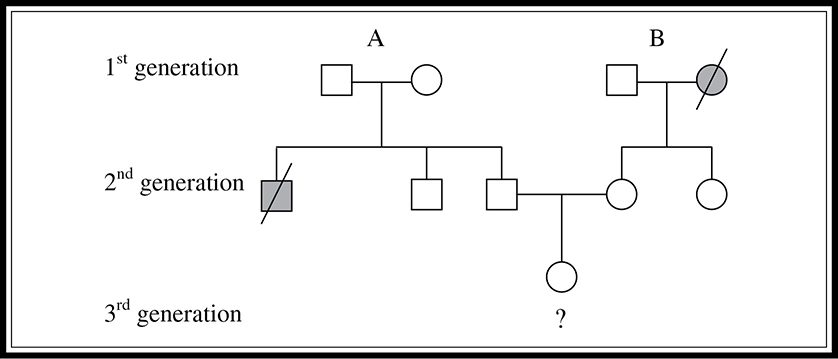
Figure 9.9 Three-generation pedigree indicating probability of inheriting a particular disease.
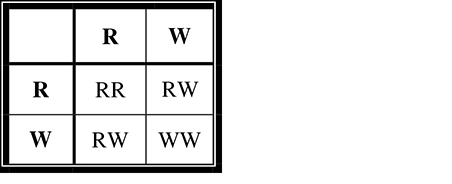
First, we can determine the father’s (second-generation) probability of being a carrier. We know that both of his parents must be carriers with a genotype of Dd. Why is this the case? Although neither parent has the condition, they must both be carriers for his brother to have received two recessive alleles and thus have contracted the disease. How can we calculate the potential probability of the father being a carrier? We construct a Punnett square for a monohybrid cross of the father’s parents (first generation) (Figure 9.10.
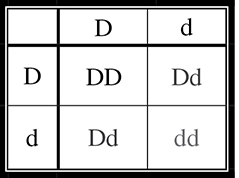
Figure 9.10 A Punnett square.
We know with certainty that he is not dd, otherwise, he would have the condition. This leaves three equally likely possible genotypes for the father, two of which are “carrier” genotypes (Dd). Thus, the probability of his being a carrier is ⅔.
What is the probability that the mother (second generation) is a carrier? We don’t even need a Punnett square to determine this one. Her mother (first generation) died of the condition, which means that she must have been dd, and thus must have passed along a d to each of her children. The mother in question does not have the condition, so she must have a D as well. Therefore her genotype must be Dd.
To determine the probability that both parents are carriers, apply the law of multiplication with probabilities (similar to tossing a coin) and use the following formula:
![]()
(where PF, PM = probabilities of father, mother being carriers).
Now that we have determined the probability that they are both carriers, we need to determine the probability that one of their offspring will have the condition. Their Punnett square would be the same as that shown in Figure 9.10, and we can see that the probability of having a child with the recessive condition is ¼. Again, we use the law of multiplication and see that the probability of this couple having a child with the condition is ⅔ × ¼ = 1⁄6.
If these two second-generation parents had a child with the recessive condition, what would the probability be of their next child having the condition? It would no longer be 1⁄6; once they have had a child with the condition, we would know with 100 percent certainty that they are heterozygous carriers. Thus, the probability that their next child will have the condition is ¼, as shown in Figure 9.10.
Common Disorders
There are many simple recessive disorders in which a person must be homozygous recessive for the gene in question to have the disease. Some of the most common examples are Tay-Sachs disease, cystic fibrosis, sickle cell anemia, phenylketonuria, and albinism. These diseases are commonly used as examples on the AP Biology exam and could also aid you in constructing a well-supported essay answer to a question about heredity and inherited disorders.
Tay-Sachs disease is a fatal genetic disorder that renders the body unable to break down a particular type of lipid that accumulates in the brain and eventually causes blindness and brain damage. Individuals with this disease typically do not survive more than a few years. Carriers of this disease do not show any of the effects of the disease, and thus the allele is preserved in the population because carriers usually live to reproduce and potentially pass on the recessive copy of the allele. This disease is found in a higher-than-normal percentage of people of eastern European Jewish descent.
Cystic fibrosis (CF), a recessive disorder, is the most common fatal genetic disease in this country. The gene for this disease is located on chromosome 7. The normal allele for this gene is involved in cellular chloride ion transport. A defective version of this gene results in the excessive secretion of a thick mucus, which accumulates in the lungs and digestive tract. Left untreated, children with CF die at a very young age. Statistically, 1 in 25 Caucasians is a carrier for this disease.
Sickle cell anemia is a common recessive disease that occurs as a result of an improper amino acid substitution during translation of an important red blood cell protein called hemoglobin. It results in the formation of a hemoglobin protein that is less efficient at carrying oxygen. It also causes hemoglobin to deform to a sickle shape when the oxygen content of the blood is low, causing pain, muscle weakness, and fatigue.
Sickle cell anemia is the most common inherited disease among African Americans. It affects 1 out of every 400 African Americans, and 1 out of 10 African Americans is a carrier of the disease. The recessive trait is so prevalent because carriers (who are said to have sickle cell “trait”) have increased resistance to malaria. In tropical regions, where malaria occurs, the sickle cell trait actually increases an individual’s probability of survival, and thus the trait’s presence in the population increases (heterozygote advantage).
Phenylketonuria (PKU) is another autosomal recessive disease caused by a single gene defect. Children with PKU are unable to successfully digest phenylalanine (an amino acid). This leads to the accumulation of a by-product in the blood that can cause mental retardation. If the disease is caught early, retardation can be prevented by avoiding phenylalanine in the diet.
Dominant disorders are less common in humans. One example of a dominant disorder is Huntington disease, a fatal disease that causes the breakdown of the nervous system. It does not show itself until a person is in their 30s or 40s, and individuals afflicted with this condition have a 50 percent chance of passing it to their offspring.
Why are lethal dominant alleles less common than lethal recessive alleles? Think about how recessive alleles often are passed on from generation to generation. An individual can be a carrier of a recessive condition and pass it along without even knowing it. On the other hand, it is impossible to be an unaffected carrier of a dominant condition, and many lethal conditions have unfortunately killed the individual before reproductive maturity has been achieved. This makes it more difficult for the dominant gene to be passed along. To remain prevalent in the population, a dominant disorder must not kill the individual until reproduction has occurred.
Chromosomal Complications
We have spent a lot of time discussing how genes are inherited and passed from generation to generation. It is also important to discuss the situations in which something goes wrong with the chromosomes themselves that affects the inheritance of genes by the offspring. Nondisjunction is an error in homologous chromosome separation. It can occur during meiosis I or II. The result is that one gamete receives too many of one kind of chromosome, and another gamete receives none of a particular chromosome. The fusing of an abnormal gamete with a normal one can lead to the production of offspring with an abnormal number of chromosomes (aneuploidy).
Down syndrome is a classic aneuploid example, affecting 1 out of every 700 children born in this country. It most often involves a trisomy of chromosome 21, and leads to mental retardation, heart defects, short stature, and characteristic facial features. Most people with trisomy 21 are sterile.
Trisomy 21 is not the only form of nondisjunction caused by error in the chromosome separation process. Trisomy 13, also known as Patau syndrome, causes serious brain and circulatory defects. Trisomy 18, also known as Edwards syndrome, can affect all organs. It is rare for a baby to survive for more than a year with either of these two conditions. There are also syndromes involving aneuploidy of the sex chromosomes. Males can receive an extra Y chromosome (XYY). Although this nondisjunction does not seem to produce a major syndrome, XYY males tend to be taller than average, and some geneticists believe they display a higher degree of aggressive behavior. A male can receive an extra X chromosome, as in Klinefelter syndrome (XXY). These infertile individuals have male sex organs but show several feminine body characteristics. Nondisjunction occurs in females as well. Females who are XXX have no real syndrome. Females who are missing an X chromosome (XO) have a condition called Turner syndrome. XO individuals are sterile females who possess sex organs that fail to mature at puberty.
Trisomies are not the only kind of chromosomal abnormalities that lead to inherited diseases. A deletion occurs when a piece of the chromosome is lost in the developmental process. Deletions, such as cri-du-chat syndrome, can lead to problems. This syndrome occurs with a deletion in chromosome 5 that leads to mental retardation, abnormal facial features, and a small head. Most affected individuals die very young.
Chromosomal translocations, in which a piece of one chromosome is attached to another, nonhomologous chromosome, can cause major problems. Chronic myelogenous leukemia is a cancer affecting white blood cell precursor cells. In this disease, a portion of chromosome 22 has been swapped with a piece of chromosome 9.
A chromosome inversion occurs when a portion of a chromosome separates and reattaches in the opposite direction. This can have no effect at all, or it can render a gene nonfunctional if it occurs in the middle of a sequence. A chromosome duplication results in the repetition of a genetic segment. A chromosome duplication results in the repetition of a genetic segment . . . whoops . . . sorry. . . . Duplications often have serious effects on an organism.
These are the major concepts of heredity with which the AP Biology exam writers would like you to be familiar. Try the practice problems that follow and be sure you are able to construct, read, and analyze both Punnett squares and pedigrees, keeping in mind the laws of probability.
![]() Review Questions
Review Questions
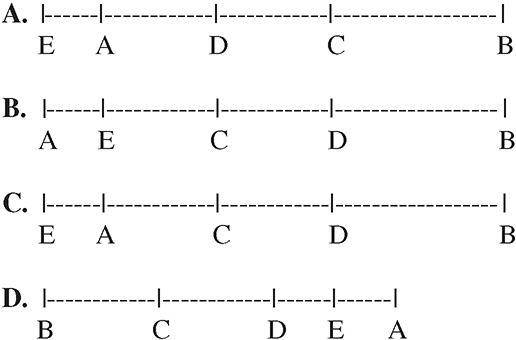
1. The following crossover frequencies were noted via experimentation for a set of five genes on a single chromosome:
A and B → 35%
B and C → 15%
A and C → 20%
A and D → 10%
D and B → 25%
A and E → 5%
B and E → 40%
Pick the answer that most likely represents the relative positions of the five genes.
2. Imagine that in squirrels, gray color (G) is dominant over black color (g). A black squirrel has the genotype gg. Crossing a gray squirrel with which of the following would let you know with the most certainty the genotype of the gray squirrel?
A. GG
B. Gg
C. gg
D. Cannot be determined from the information given
3. From a cross of AABbCC with AaBbCc, what is the probability that the offspring will display a genotype of AaBbCc?
A. ½
B. ⅓
C. ¼
D. ⅛
Use the following pedigree of an autosomal recessive condition for questions 4—6.
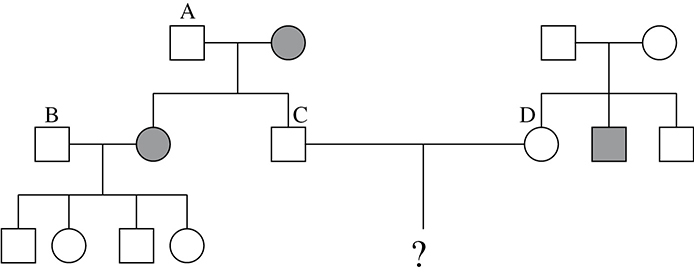
4. What is the genotype of person A?
A. Bb
B. BB
C. bb
D. Cannot be determined from the giveninformation
5. What is the most likely genotype of person B?
A. Bb
B. BB
C. bb
D. Cannot be determined from the information given
6. What is the probability that persons C and D would have a child with the condition?
A. ½
B. ¼
C. 1⁄6
D. ⅛
7. Which of the following disorders is X-linked?
A. Tay-Sachs disease
B. Cystic fibrosis
C. Hemophilia
D. Albinism
8. A court case is trying to determine the father of a particular baby. The mother has type O blood, and the baby has type B blood. Which of the following blood types would mean that the man was definitely not the father of the baby?
A. B and A
B. AB and A
C. O and B
D. O and A
9. Assume that gray squirrel color results from a dominant allele G. The father squirrel is black, the mother squirrel is gray, and their first baby is black. What is the probability that their second baby is also black?
A. 1.00
B. 0.75
C. 0.50
D. 0.25
10. Imagine that tulips are either yellow or white. You start growing tulips and find out that if you want to get yellow tulips, then at least one of the parents must be yellow. Which color is dominant?
A. White
B. Yellow
C. Neither; it is some form of intermediate inheritance
D. Cannot be determined from the given information
11. Suppose that 200 red snapdragons were mated with 200 white snapdragons and they produced only pink snapdragons. The mating of two pink snapdragons would most likely result in offspring that are
A. 50 percent pink, 25 percent red, 25 percent white
B. 100 percent pink
C. 25 percent pink, 50 percent red, 25 percent white
D. 75 percent red, 25 percent white
12. Which of the following represents the number of possible gametes produced from a genotype of RrBBCcDDEe?
A. 2
B. 4
C. 8
D. 16
13. Which of the following diseases is not caused by trisomy nondisjunction?
A. Down syndrome
B. Klinefelter syndrome
C. Turner syndrome
D. Patau syndrome
14. The pedigree below is most likely a pedigree of a condition of which type of inheritance?
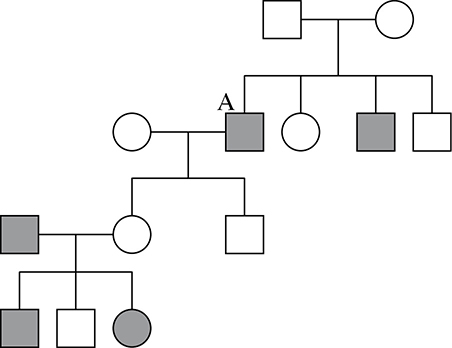
A. Autosomal dominant
B. Autosomal recessive
C. Sex-linked dominant
D. Sex-linked recessive
![]() Answers and Explanations
Answers and Explanations
1. A—The crossover frequencies are an indication of the distance between the different genes on a chromosome. The farther apart they are, the greater chance there is that they will cross over during prophase I of meiosis. You are first told that A and B cross over with a frequency of 35 percent, so imagine that they are 35 units apart on a chromosome map.
![]()
We can then tell you that B and C have a frequency of 15 percent. They are 15 units apart on the map, but you cannot yet be sure what side of gene A that C is on. Gene A and C cross with 20 percent frequency. This means that gene C must be in between A and B.
![]()
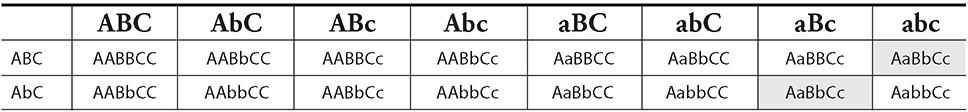
Gene A crosses over with D 10 percent of the time, and D crosses with B 25 percent of the time; therefore, D must also be in between A and B. It is closer to A than it is to B. You can use this knowledge to eliminate answer choices B and C.
![]()
Gene A crosses over with E with a frequency of 5 percent. You do not know which side of A gene E is on until you know its crossover frequency with B. Because the question tells you that it has a 40 percent frequency with B, you know that it must be on the left of A. This completes your map, leaving A as the correct answer.
2. C—This is a test cross. To determine the genotype of an individual showing the dominant phenotype, you cross that individual with a homozygous recessive individual for the same trait. If they have no offspring with the recessive phenotype, then the individual displaying the dominant phenotype is most likely GG. If approximately one-half of the offspring have the recessive phenotype, you know the individual has the genotype Gg.
3. D—The Punnett square shown below shows all the possible gamete combinations from this cross. Two-sixteenths or one-eighth of the possible gametes will be AaBbCc. A quick way to determine the number of possible gametes that an individual can produce given a certain genotype is to use the formula 2n. For example, an individual who is AABbCc can have 22 = 4 possible gametes because Bb and Cc are heterozygous.
4. A—Person A must have genotype Bb because he has some children that have the recessive condition and some that do not. Because his wife is pure recessive, she can contribute only a b. The father must therefore be the one who contributes the B to the child who does not have the condition, and the second b to the one with the condition.
5. B—Person B most likely has a genotype of BB. Because he does not have the condition, we know that his genotype is either BB or Bb. If it were Bb, then when crossed with his wife who has a genotype of bb, 50 percent of his children would be expected to have the recessive condition. None of the children have the condition, which leads you to believe that he is most likely BB. (This test is, of course, not 100 percent accurate. Answer choice B is not certain, but is the most probable conclusion.)
6. C—We know that neither parent in the question has the recessive condition. We therefore need to calculate the probability that each of them is Bb. The probability that person C is Bb is 1. Because his mother has the condition, she must pass a b to him during gamete formation. So the only possible genotypes he can have are Bb and bb. Since he does not have the condition, he must be Bb with a probability of 1. The probability that person D is Bb is 0.67. Neither of her parents has the condition, but she has a brother who is bb. This means that each of her parents must be a carrier for the condition (Bb). You know that this woman is not bb, because she does not have the condition. As a result, there are only three possible genotypes from the cross remaining. Two of these three are Bb, giving her a probability of ⅔, or 0.67, of being Bb. The probability that both person C and person D are Bb is (1) × (0.67) = (0.67). Now it is necessary to calculate the probability that two Bb parents will produce a kid who is bb. The Punnett square says that there is a 0.25 chance of this result. To calculate the probability that they will have a child with the recessive condition, you multiply the probability that they are both Bb (0.67) times the probability that two individuals Bb will produce a bb child (0.25). Thus, the probability of an affected child being produced from these two parents is 1⁄6.
7. C—Hemophilia is an X-linked condition. An XY male with hemophilia gets his Y chromosome from his father, and his X chromosome from his mother. All that is needed for the hemophilia condition to occur is a copy of the defective recessive allele from his mother.
8. D—Types O and A would prove that he was not the father of this particular child. If the mother has type O blood, this means that her genotype is ii and she must pass along an i allele to her child. The baby has type B blood, and her genotype could be IBi or IBIB. Since the mother must give an i, then the baby’s genotype must be IBi. It follows that the father must provide the IB allele to the baby to complete the known genotype. If he is type O, he won’t have an IB to pass along since his genotype would be ii. This would also be the case if he were type A, because his genotype would be either IAIA or IAi. Therefore, those two blood types would prove that he is not the father of this child.
9. C—To figure out this problem, you need to know the genotype of the mother. The father is black, meaning that his genotype is gg. The two of them produced a squirrel that is also black, which means that the gray mother gave a g to the baby. The mother’s genotype is Gg. A cross of Gg × gg produces a phenotype ratio of 1:1 gray:black. They have a 0.5 chance of producing another black baby.
10. B—According to this scenario, yellow and white are the only colors possible. If white were dominant, and both parents were Ww, you could produce a yellow offspring if the two recessive w’s combined. If it were intermediate inheritance, you probably would not produce a straight yellow tulip in the offspring because they would either meet halfway (incomplete dominance), or both express fully (codominance). If yellow were dominant, then you could produce a yellow offspring only if there were a Y allele in one of the parents. A cross of yy × yy would produce only white tulips if white were recessive.
11. A—This problem involves incomplete dominance. The genotype of the pink offspring from the first generation is RW. When the two RW snapdragons are mated together, they produce the following results:
The offspring will be 25 percent red (RR), 50 percent pink (RW), and 25 percent white (WW).
12. C—In a problem like this, you will save time by thinking about the laws of probability. The genotype is RrBBCcDDEe. How many possible combinations of the R gene are there? There are two: R and r. How many for B? Only one: B. Following the same logic, C has two and D has one. Now you multiply the possibilities: (2 × 1 × 2 × 1 × 2) = 8. There are 8 possible gametes from this genotype. Another way to arrive at this answer is by use of the expression 2n, where n is the number of hybrid traits being examined. In this case, it would be 23 or 8 possible gametes.
13. C—Down syndrome is most often due to a trisomy of chromosome 21. Klinefelter syndrome is a trisomy of the sex chromosomes (XXY). Patau syndrome is a trisomy of chromosome 15. Edwards syndrome is a trisomy of chromosome 18. Turner syndrome, the only nontrisomy listed in this problem, is a monosomy of the sex chromosomes (XO).
14. D—This is most likely a sex-linked recessive disease. The father in the first generation does not have the condition, so his genotype would be XNY. The original couple has four children, two boys with the condition, and one girl and one boy without the condition. The genotype of the boys with the condition would be XnY. This means that the original mother’s genotype would be XNXn—thus she is a carrier. One of the children who inherited the condition has children with a woman from a different family, and neither of their two children displays the condition. However, the daughter of son A has three children with a man who is XnY, and she has a daughter and a son who show the recessive condition and one normal son. This means that the daughter of son A is most likely XNXn—another carrier of the condition. This disease is a condition that is, according to the pedigree, more often seen in men, and passed along to men by the X chromosome from the mother. However, it is important to note that if a father who has the X-linked condition has a child with a female carrier for the condition, that couple can indeed produce a female with the condition.
![]() Rapid Review
Rapid Review
You should be familiar with the following terms:
Character: heritable feature, such as flower color.
Monohybrid cross: cross involving one character (Bb × Bb) → (3:1 phenotype ratio).
Dihybrid cross: cross involving two different characters (BbRr × BbRr) → (9:3:3:1 phenotype ratio).
Law of segregation: the two alleles for a trait separate during the formation of gametes—one to each gamete.
Law of independent assortment: inheritance of one trait does not interfere with the inheritance of another trait.
Law of dominance: if two opposite pure-breeding varieties (BB × bb) are crossed, all offspring resemble BB parent.
Intermediate inheritance: heterozygous (Yy) individual shows characteristics unlike either parent.
✵ Incomplete dominance: Yy produces an intermediate phenotype between YY and yy (snapdragons).
✵ Codominance: both alleles express themselves fully in a Yy individual—(MN blood groups).
Polygenic traits: traits that are affected by more than one gene (eye color, skin color).
Multiple alleles: traits that correspond to more than two alleles (ABO blood type: IA, IB, i).
Epistasis: a gene at one locus alters the phenotypic expression of a gene at another locus (coat color in mice).
Pleiotropy: a single gene has multiple effects on an organism (sickle cell anemia).
Sex determination: males are XY, females are XX.
Autosomal chromosome: not involved in gender.
Fruit flies: wild-type traits are the normal phenotype; mutant traits are those that are different from normal.
Sex-linked traits: passed along the X chromosome; more common in males than females (males have only one X) (e.g., hemophilia [can’t clot blood], Duchenne’s muscular dystrophy [muscle weakness], colorblindness).
X inactivation: one of two X chromosomes is randomly inactivated and remains coiled as a Barr body.
Holandric trait: one that is inherited via the Y chromosome.
Linked genes: genes that lie along the same chromosome and do not follow the law of independent assortment.
✵ Crossover: a form of genetic recombination that occurs during prophase I of meiosis.
✵ The further apart two genes are along a chromosome, the more often they will cross over.
Linkage map: genetic map put together using crossover frequencies.
✵ Can determine the relative location of a set of genes according to how often they cross over.
✵ If two genes cross over in 20 percent of the crosses, they are 20 map units apart, etc.
Law of multiplication: To determine the probability that two random events will occur in succession, multiply the probability of the first event by the probability of the second event. (Useful in pedigree analysis!)
Pedigree: family tree used to describe genetic relationships (use pedigree diagram in review question 14 for clearer understanding). To calculate the risk a couple faces of having a child that has a recessive (bb) condition, first determine the probability that both parents are Bb (if neither have the condition), or the probability that one is Bb (if one has the condition). Once determined, multiply this probability times the probability that a Bb × Bb cross will produce a bb (¼) or that a bb × Bb will produce a bb (½).
Autosomal Recessive Disorders
Tay-Sachs: fatal, storage disease, lipid builds up in brain, mental retardation, increased incidence in eastern European Jews.
Cystic fibrosis: increased mucus buildup in lungs; untreated children die at young age; 1 in 25 Caucasians is a carrier.
Sickle cell anemia: caused by error of single amino acid; hemoglobin is less able to carry O2, and sickles when O2 content of blood is low; 1 in 10 African Americans is a carrier. Heterozygous condition protects against malaria.
Phenylketonuria: inability to digest phenylalanine, which can cause mental retardation if not avoided in diet.
Autosomal dominant disorders: Huntington disease (nervous system disease) and achondroplasia (dwarfism).
Nondisjunction: error in which homologous chromosomes do not separate properly.
✵ Monosomy: (one copy): Turner syndrome.
✵ Trisomy: (three copies): Down syndrome (21), Patau syndrome (13), Edwards syndrome (18).
Klinefelter syndrome: XXY; XYY males, XXX females.
Chromosome disorders: deletion (cri-du-chat), inversions, duplications, and translocations (leukemia).