5 Steps to a 5: AP Biology - Mark Anestis 2021
STEP 4 Review the Knowledge You Need to Score High
CHAPTER 11 Evolution
Exam Weight: 13—20%
IN THIS CHAPTER
Summary: This chapter discusses evolution and the four major modes in which it occurs. It introduces you to the various forms of selection: natural, directional, stabilizing, disruptive, sexual, and artificial. It discusses the two main forms of speciation (allopatric and sympatric) and briefly touches on the theory behind how life on this planet emerged many years ago.

Key Ideas
![]() The four major modes of evolution are genetic drift, gene flow, mutation, and natural selection.
The four major modes of evolution are genetic drift, gene flow, mutation, and natural selection.
![]() Natural selection is based on three conditions: variation, heritability, and differential reproductive success.
Natural selection is based on three conditions: variation, heritability, and differential reproductive success.
![]() There are four basic patterns of evolution: co-evolution, convergent evolution, divergent evolution, and parallel evolution.
There are four basic patterns of evolution: co-evolution, convergent evolution, divergent evolution, and parallel evolution.
![]() Sources of variation within populations: mutation, sexual reproduction, and balanced polymorphism.
Sources of variation within populations: mutation, sexual reproduction, and balanced polymorphism.
![]() Hardy-Weinberg conditions: no mutations, no gene flow, no genetic drift, no natural selection, and random mating.
Hardy-Weinberg conditions: no mutations, no gene flow, no genetic drift, no natural selection, and random mating.
![]() Hardy-Weinberg equations: p + q = 1 and p2 + 2pq + q2 = 1.
Hardy-Weinberg equations: p + q = 1 and p2 + 2pq + q2 = 1.
![]() Evidence for evolution: homologous characters, embryology, and vestigial structures.
Evidence for evolution: homologous characters, embryology, and vestigial structures.
![]() Evolutionary relationships are shown with phylogenetic trees and/or cladograms using fossils, molecular clocks, and/or molecular data.
Evolutionary relationships are shown with phylogenetic trees and/or cladograms using fossils, molecular clocks, and/or molecular data.
Introduction
This chapter begins with an introduction to the concept of evolution and the four major modes in which it occurs. From there we focus more closely on natural selection and the work of Lamarck and Darwin. We then briefly touch on adaptations before looking at the various types of selection: directional, stabilizing, disruptive, sexual, and artificial selection. This is followed by a quick look at the sources of variation within populations followed by a look at the two main types of speciation: allopatric and sympatric. Next, will come the yucky math portion of the chapter: the Hardy-Weinberg equation and the conditions necessary for its existence. The chapter concludes with a look at the existing evidence in support of the theory of evolution and a discussion of how life on this planet emerged so many years ago.
Definition of Evolution
How often have you heard executives report that “the idea evolved into a successful proj-ect” or popular science show narrators describe how a star “has been evolving for millions of years”? Evolution is no longer strictly a biological term since every academic field and nonacademic industry uses it. Such uses of the verb evolve reveal its meaning in its simplest form—to evolve means to change. For the AP Biology exam, however, you should remember the biological definition of evolution: descent with modification. Don’t let the general uses of the word mislead you; a key part of this definition is descent, which can happen only when one group of organisms gives rise to another. When you see the word evolution, think of something that happens in populations, not in individuals.
More specifically, evolution describes change in allele frequencies in populations over time. When one generation of organisms (whether algae or giraffes or ferns) reproduces and creates the next, the frequencies of the alleles for the various genes represented in the population may be different from what they were in the parent generation. Frequencies can change so much that certain alleles are lost or others become fixed—all individuals have the same allele for that character. Over many generations, the species can change so much that it becomes quite different from the ancestral species, or a part of the population can branch off and become a new species (speciation). Why do we see this change in allele frequencies with time?
EVO-1
Evolution is characterized by a change in the genetic makeup of a population over time and is supported by multiple lines of evidence.
Allele frequencies may change because of random factors or by natural selection. Let’s consider chance events first. Imagine a population of fish in a large pond that exhibits two alleles for fin length (short and long) and is isolated from other populations of the same species. One day, a tornado kills 50 percent of the fish population. Completely by chance, most of the fish killed possess the long-fin allele, and very few of these individuals are left in the population. In the next generation, there are many fewer fish with long fins because fewer long-finned fish were left to reproduce; that allele is much more poorly represented in the pond than it was in the original parent generation before the catastrophe. This is an example of genetic drift (Figure 11.1: a change in allele frequencies that is due to chance events. When drift dramatically reduces population size, we call it a bottleneck.
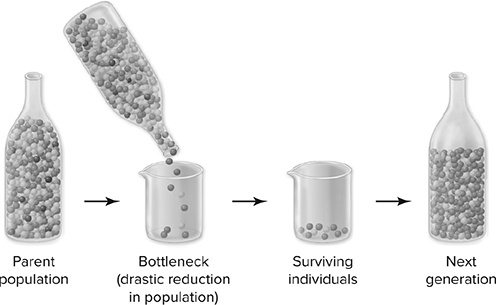
Figure 11.1 Genetic drift: A bottleneck effect. The parent population contains roughly equal numbers of green and yellow individuals and a small number of red -individuals. By chance, the few remaining individuals that contribute to the next -generation are mostly green. The bottleneck occurs because so few individuals form the next generation, as might happen after an epidemic or a catastrophic storm. (Reproduced with permission from Raven P, Johnson G, Mason K, Losos J, Duncan T; Biology, 12th ed. New York: McGraw Hill; 2020)
Now imagine that the same pond becomes connected to another pond by a small stream. The two populations mix, and by chance, all the long-finned fish migrate to the other pond, and no long-finned fish migrate in. Again, which individuals migrated was random in this example; thus, there will be a change in the allele frequencies in the next generation. This is an example of gene flow, or the change in allele frequencies as genes from one population are incorporated into another.
Gene flow (also more loosely known as migration when the individuals are actively relocating) is random with respect to which organisms succeed, but keep in mind that we could think of situations in which migration is not random. For example, if only the short-finned fish could fit in the stream connecting the two ponds, the alleles represented in the subsequent generation would not be random with respect to that allele. We also have not stated that the short-finned fish have an advantage by swimming to the other pond—if they did, this would be an example of natural selection, which we’ll discuss below.
Finally, let’s consider mutation, the third random event that can cause changes in allele frequencies. Mutation is always random with respect to which genes are affected, although the changes in allele frequencies that occur as a result of the mutation may not be. Let’s say that a mutation occurs in the offspring of a fish in our hypothetical pond. The mutation creates a new allele. As a result, the allele frequencies in the offspring generation has changed, simply because we have added a new allele (remember that allele frequencies for a given gene always add up to one). As you can imagine, one mutation on its own does not have the potential to dramatically alter the allele frequencies in a population, unless this is a really small pond! But mutation is extremely important because it is the basis of the variation we see in the first place and it is a very strong force when it is paired with natural selection.
The five major modes of evolution (Figure 11.2 are
1. Genetic drift
2. Gene flow (also called migration)
3. Mutation
4. Natural selection
5. Nonrandom mating
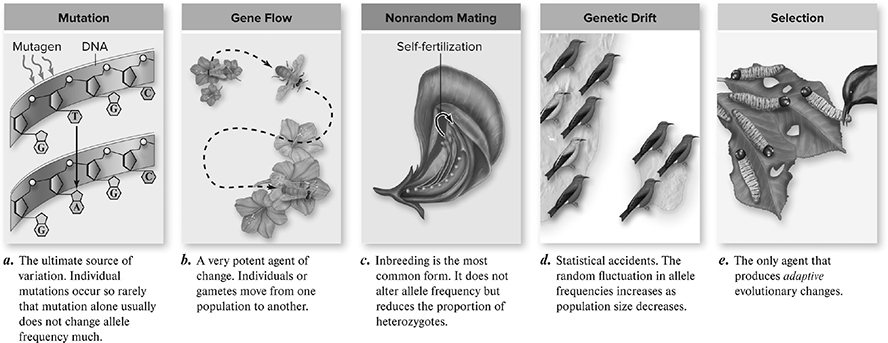
Figure 11.2 Five agents of evolutionary change. (Reproduced with permission from Raven P, Johnson G, Mason K, Losos J, Duncan T; Biology, 12th ed. New York: McGraw Hill; 2020)
Nonrandom mating or sexual selection is considered by some to be a subset of natural selection, while others believe that it is separate from natural selection. In random mating, organisms participate in intrasexual selection, which represent competitive interactions between the same sex (male-to-male or female-to-female) and intersexual selection, which represents the selection of reproductive partners of the opposite sex. This leads to the evolution of secondary sexual characteristics for organisms to persuade members of the opposite sex such as the female peahen preferring a male with the greatest number of eyespots on his feathers (Figure 11.3.

Figure 11.3 Products of sexual selection. In bird species such as the peacock, Pavo cristatus, males use their much longer tail feathers in courtship displays. (Reproduced with permission from Raven P, Johnson G, Mason K, Losos J, Duncan T; Biology, 12th ed. New York: McGraw Hill; 2020)
Remember that the first three factors act randomly with respect to the alleles in the population—which alleles increase and which decrease in frequency are determined by chance events, not because some alleles are inherently better than others. We’ll now turn to the fourth mode or process of evolution, natural selection, where the modification that occurs with descent is nonrandom.
Natural Selection
EVO-1
Evolution is characterized by a change in the genetic makeup of a population over time and is supported by multiple lines of evidence.
Probably the biggest mistake people make when thinking about natural selection is thinking that it is synonymous with evolution. Natural selection is only one process by which evolution occurs (the others are discussed in the previous section). However, it is an important process because it has been instrumental in shaping the natural world. Because of the theory of natural selection, we can explain why organisms look and behave the way they do.
Natural selection is based on three conditions:
1. Variation: for natural selection to occur, a population must exhibit phenotypic variance— in other words, differences must exist between individuals, even if they are slight.
2. Heritability: parents must be able to pass on the traits that are under natural selection. If a trait cannot be inherited, it cannot be selected for or against.
3. Differential reproductive success: this sounds complicated, but it’s a simple concept. Reproductive success measures how many offspring you produce that survive relative to how many the other individuals in your population produce. The condition simply states that there must be variation between parents in how many offspring they produce as a result of the different traits that the parents have.
It is easiest to illustrate natural selection with an example. Let’s revisit our pond before the tornado came, where short- and long-finned fish inhabit murky waters. A new predator invades the pond. Fin length determines swimming speed (longer fins allow a fish to swim faster), and only the fastest fish can escape the predator. How would you expect the allele frequencies to change under these conditions? Fish with what length fin would be eaten the most? Because the short-finned fish would be the slowest, they would be featured on the menu. But the long-finned fish, able to escape this new predator, would survive and reproduce, and the frequency of the long-fin allele would increase relative to the short-fin allele. We have created a situation in which allele frequencies change as a result of a nonrandom event; the predator’s presence results in a predictable decrease in the short-fin allele and a consequent increase in the long-fin allele. Remember that allele frequencies always add up to 100 percent, so the long-finned fish don’t have to do particularly well for the long-fin allele to increase—they only have to do well relative to the short-finned fish. The actual numbers of fish could decrease for both variants of this fish species.
Why aren’t organisms perfectly adapted to their environments? Since natural selection increases the frequencies of advantageous alleles, why don’t we get to a point where all individuals have all the best alleles? For one, different alleles confer different ad-vantages in different environments. Furthermore, remember that the environment—which includes everything from habitat, to climate, to competitors, to predators, to food resources—is constantly changing. Species are therefore also constantly changing as the traits that give them an advantage also change. In cases where a trait becomes unconditionally advantageous, we do in fact see fixed alleles; for example, all spiders have eight legs because the alternatives just aren’t as good under any circumstances. But where there are heritable characters that both vary and confer fitness advantages (or disadvantages) on their host organisms, natural selection can occur.
Lamarck and Darwin
The two key figures whose research you should know for the evolution section of the AP Biology exam are Jean-Baptiste Lamarck and Charles Darwin. Lamarck proposed the idea that evolution occurs by the inheritance of acquired characters. The classic example is giraffe necks: Lamarck proposed that giraffes evolved long necks because individuals were constantly reaching for the leaves at the tops of trees. A giraffe’s neck lengthened during its lifetime, and then that giraffe’s offspring had a long neck because of all that straining its parents did. The key here is that change happened within organisms during their lifetimes and then the change in the trait was passed on.
What’s wrong with Lamarck’s theory? Try explaining to yourself how the changed character could be passed on to the offspring. The answer is that it couldn’t—the instructions in the sex chromosomes that direct the production of offspring cannot be changed after they are created at the birth of an organism. Lamarck confused genetic and environmental (postconceptive) change, which is not surprising because no one had discovered genes yet.
Darwin had another idea, one that ended up being entirely consistent with Mendelian genetics (although Mendel had already written his thesis during Darwin’s time, it is rumored that his book sat on Darwin’s shelf, with the pages still uncut, until Darwin’s death). Darwin suggested the idea of natural selection described above and coined the phrase “survival of the fittest.” Although he didn’t call them genes, he proposed a hypothetical unit of heredity that passed from parent to offspring. Incidentally, a man named Wallace also came up with the idea of natural selection during the same time, but Darwin got the publication out first and has become famous as a result.
Adaptations
An adaptation is a trait that if altered, affects the fitness of the organism. Adaptations are the result of natural selection and can include not only physical traits such as eyes, fingernails, and livers but also the intangible traits of organisms. For example, lifespan length is an adaptation, albeit a variable one. Mating behavior is also an adaptation—it has been selected by natural selection because it is an effective strategy. An individual with a different form of mating behavior may do better or worse than the average, but a change is likely to have some effect on reproductive success. For example, individuals whose mating strategy is to attempt to court women by running at them, arms flailing while screaming wildly, and salivating heavily, do worse than the average male.
Let’s take a look at how such a behavioral adaptation can evolve. Reproductive maturity is a good example. Female chimpanzees become reproductively mature at around the age of 13. Females that mature at age 12 spend less time growing and may therefore be more susceptible to problems with pregnancy. Females that mature at 14 have lost valuable time—their earlier-maturing peers have gained a year on them. You can imagine that from generation to generation, females that matured at age 13 became better represented in the population compared to faster and slower maturers. Although there will always be individuals that differ from the mode, we can view age at reproductive maturation as an adaptation.
Types of Selection
Natural selection can change the frequencies of alleles in populations through various processes. The most commonly described are the following three (Figure 11.4:
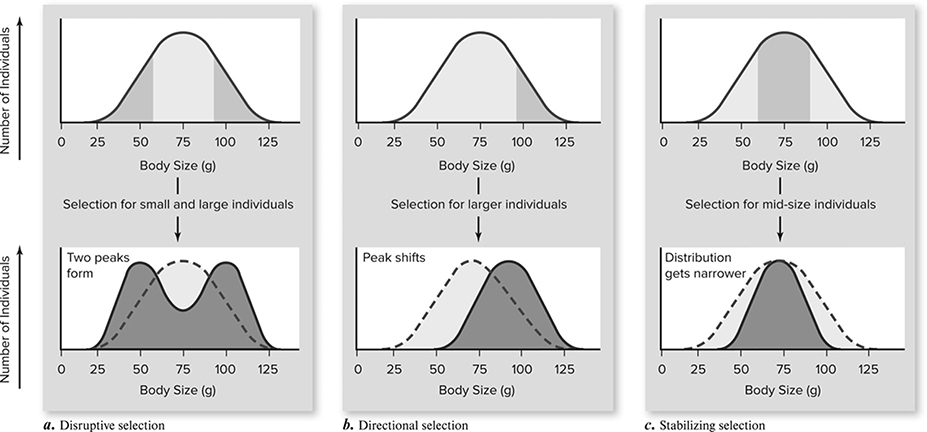
Figure 11.4 Three types of selection: (a) disruptive; (b) directional; (c) stabilizing. (Reproduced with permission from Raven P, Johnson G, Mason K, Losos J, Duncan T; Biology, 12th ed. New York: McGraw Hill; 2020)
a. Directional selection. This occurs when members of a population at one end of a spectrum are selected against, while those at the other end are selected for. For example, imagine a population of elephants with various-sized trunks. In this particular environment, much more food is available in the very tall trees than in the shorter trees. Elephants with what length trunk will survive and reproduce the most successfully? Those with the longest trunks. Those with shorter trunks will be strongly selected against (and those in the middle will also be in the middle in terms of success). Over time we expect to see an increasing percentage of elephants with long trunks (how quickly this change occurs depends on the strength of selection—if all the short-trunked elephants die, we can imagine that the allele frequencies will change very quickly).
b. Stabilizing selection. This describes selection for the mean of a population for a given allele. A real example of this is human infant birth weight—it is a disadvantage to be really small or really big, and it is best to be somewhere in between. Stabilizing selection has the effect of reducing variation in a population.
c. Disruptive selection. Also known as diversifying selection, this process can be regarded as being the opposite of stabilizing selection. We say that selection is disruptive when individuals at the two extremes of a spectrum of variation do better than the more common forms in the middle. Snail shell color is an example of disruptive selection. Imagine an environment in which snails with very dark shells and those with very light shells are best able to hide from predators. Those with an in-between shell color are gulped up like escargot at a cocktail party, creating the double-hump curve.
Mike (freshman in college): “Learn these selection types . . . they make good multiple-choice questions.”
EVO-3
Life continues to evolve within a changing environment.
These three processes describe the way in which allele frequencies can change as a result of the forces of natural selection. It is also important to remember two other types of selection that complement natural selection: sexual selection and artificial selection.
Sexual selection occurs because individuals differ in mating success. In other words, because not all individuals will have the maximum number of possible offspring, there must be some reason why some individuals have greater reproductive success than others. Think about how this is different from natural selection, which includes both reproduction and survival. Sexual selection is purely about access to mating opportunities.
Sexual selection occurs by two primary processes: within-sex competition and choice. In mammals and many nonmammalian species, females are limited in the number of offspring they can produce in their lifetimes (because of internal gestation), while males are not (because sperm are cheap to produce and few males participate in offspring care). Which sex do you think will compete, and which sex will be choosier? In most mammals for instance, males compete and females choose. It makes sense that males have to compete because females are a limiting resource, and it makes sense that females are choosy because they invest a lot in each reproductive effort. This leads to the evolution of characters that are designed for two main functions: (1) as weaponry or other tools for male competition (e.g., large testes for sperm competition) and (2) as traits that increase mating opportunities because females prefer to mate with males who have them (e.g., colorful feathers in many birds).
On what do females base their choices? While you need not become an expert on this matter, it is important to remember that female mate choice for certain characters is not random. One hypothesis for why females choose males with colorful feathers, for example, is that colorful feathers indicate good genes, which is important for a female’s offspring. Bright colors are costly, so a male with brightly colored feathers is probably healthy (which may, in turn, indicate an ability to reduce parasite load, for example). We call such sexually selected traits that are the result of female choice honest indicators. Keep in mind that selecting a mate for particular features does not necessarily involve conscious thought, and in most animals never does; the female does not think, “Oh! What nice feathers. He must come from good genes.” Rather, females who choose males that display honest indicators have more surviving offspring than do females who don’t, and as a result, the “choosing males with colorful feathers” trait increases in the population.
When humans become the agents of natural selection, we describe the process as artificial selection. Instead of allowing individuals to survive and reproduce as they would without human intervention, we may specifically select certain individuals to breed while restraining others from doing so. Artificial selection has resulted in the domestication of a wide range of plant and animal species and the selection of certain traits (e.g., cattle with lean meat, flowers with particular color combinations, dogs with specific kinds of skill).
Evolution Patterns
EVO-3
Life continues to evolve within a changing environment.
There are four basic patterns of evolution:
Coevolution. The mutual evolution between two species, which is exemplified by predator— prey relationships. The prey evolves in such a way that those remaining are able to escape predator attack. Eventually, some of the predators survive that can overcome this evolutionary adaptation in the prey population. This goes back and forth, over and over.
Convergent evolution. Two unrelated species evolve in a way that makes them more similar (think of them as converging on a single point). They are both responding in the same way to some environmental challenge, and this brings them closer together. We call two characters convergent characters if they are similar in two species, even though the species do not share a common ancestor. For example, birds and insects both have wings in order to fly, despite the fact that insects are not directly related to birds.
Divergent evolution. Two related species evolve in a way that makes them less similar. Divergent evolution can lead to speciation (allopatric or sympatric).
Parallel evolution. Similar evolutionary changes occurring in two species that can be related or unrelated. They are simply responding in a similar manner to a similar environmental condition.
Sources of Variation
Remember that one of the conditions for natural selection is variation. Where does this variation within populations come from?
EVO-3
Life continues to evolve within a changing environment.
1. Mutation. We already discussed mutations as a mechanism by which evolution occurs. Random changes in the DNA of an individual can introduce new alleles into a population.
SYI-3
Naturally occurring diversity among and between components within biological systems affects interactions with the environment.
2. Sexual reproduction. The three main sources of variation in sexual reproduction are crossing over which occurs in prophase 1 of meiosis 1, random assortment during metaphase 1, and the random fertlization of gametes.
3. Balanced polymorphism. Some characters are fixed, meaning that all individuals in a species or population have them: for example, all tulips develop from bulbs. However, other characters are polymorphic, meaning that there are two or more phenotypic variants. For example, tulips come in a variety of colors. If one phenotypic variant leads to increased reproductive success, we expect directional selection to eventually eliminate all other varieties. However, we can find many examples in the natural world where variation is prominent and one allele is not uniformly better than the others. The various ways in which balanced polymorphism is maintained are presented in Figure 11.5.
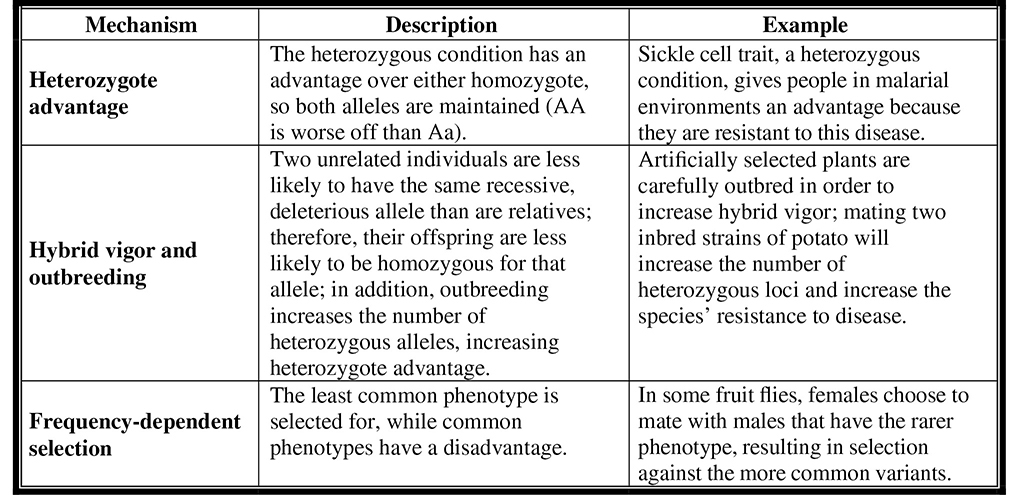
Figure 11.5 How balanced polymorphism is maintained.
Speciation
A species is a group of interbreeding (or potentially interbreeding) organisms. Speciation, the process by which new species evolve, can take one of several forms. You should be familiar with the two main forms of speciation:
1. Allopatric speciation. Interbreeding ceases because some sort of barrier separates a single population into two (an area with no food, a mountain, etc.). The two populations evolve independently (by any of the four processes discussed earlier), and if they change enough, then even if the barrier is removed, they cannot interbreed.
EVO-3
Life continues to evolve within a changing environment.
2. Sympatric speciation. Interbreeding ceases even though no physical barrier prevents it. This may take several forms.
Two other important terms are polyploidy and balanced polymorphism:
Polyploidy. A condition in which an individual has more than the normal number of sets of chromosomes. Although the individual may be healthy, it cannot reproduce with nonpolyploidic members of its species. This is unusual, but in some plants, it has resulted in new species because polyploidic individuals are only able to mate with each other.
Balanced polymorphism. This condition (described above) can also lead to speciation if two variants diverge enough to no longer be able to interbreed (if, e.g., potential mates no longer recognize each other as possible partners).
One more term to mention before moving on is adaptive radiation (Figure 11.6, which is a rapid series of speciation events that occur when one or more ancestral species invades a new environment. This process was exemplified by Darwin’s finches. If there are many ecological niches (see Chapter 12, Ecology in Further Detail), several species will evolve because each can fill a different niche.
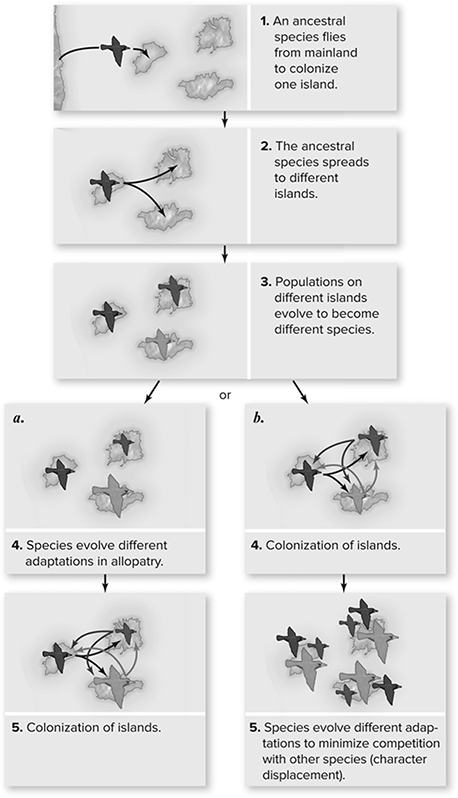
Figure 11.6 Classic model of adaptive radiation on island archipelagoes. (1) An ancestral species colonizes an island in an archipelago. Subsequently, the population colonizes other islands (2), after which the populations on the different islands speciate in allopatry (3). Then some of these new species colonize other islands, leading to local communities of two or more species. Adaptive differences can either evolve when species are in allopatry in response to different environmental conditions (a) or as the result of ecological interactions between species (b) by the process of character displacement. (Reproduced with permission from Raven P, Johnson G, Mason K, Losos J, Duncan T; Biology, 12th ed. New York: McGraw Hill; 2020)
When Evolution Is Not Occurring: Hardy-Weinberg Equilibrium
EVO-1
Evolution is characterized by a change in the genetic makeup of a population over time and is supported by multiple lines of evidence.
Evolutionary change is constantly happening in humans and other species; this seems sensible because evolution is the change in allele frequencies over time. It makes sense that these frequencies are highly variable and subject to change as the environment changes. However, biologists use a theoretical concept called the Hardy-Weinberg equilibrium to describe those special cases where a population is in stasis, or not evolving (Figure 11.7.
Only if the following conditions are met can a population be in Hardy-Weinberg equilibrium:
Hardy-Weinberg Conditions
1. No mutations
2. No gene flow
3. No genetic drift (and for this, the population must be large)
4. No natural selection (so that the traits are neutral; none gives an advantage or disadvantage)
5. Random mating
Notice items 1—4 in this list are the four modes of evolution, which makes sense—if we are trying to establish the conditions under which evolution does not occur, we must keep these processes of evolution from occurring! The fifth condition, random mating, is included because if individuals mated nonrandomly (e.g., if individuals mated with others that looked like them), the allele frequencies could change in a certain direction, and we would no longer be in equilibrium.
Determining Whether a Population Is in Hardy-Weinberg Equilibrium
Unfortunately for you, there is an equation associated with the Hardy-Weinberg equilibrium that the test writers love to put on the exam. Don’t let it scare you!
CT teacher: “Knowing how to do Hardy-Weinberg problems is worth 2 points to you . . . easy points.”
p + q = 1
This equation is used to determine if a population is in Hardy-Weinberg equilibrium. The symbol p is the frequency of allele 1 (often the dominant allele), and q is the frequency of allele 2 (often the recessive allele). Remember that the frequency of two alleles always adds up to 1 if the population is in Hardy-Weinberg equilibrium. For example, if 60 percent of the alleles for a given trait are dominant (p), then p = 0.6, and q (the recessive allele) = 1 - 0.6, or 0.4 (40 percent).
There is a second equation that goes along with this theory: p2 + 2pq + q2 = 1, where p2 and q2 represent the frequency of the two homozygous conditions (AA and aa). The frequency of the heterozygotes is pq plus qp or 2pq (Aa and aA). Since p represents the dominant allele, it makes sense that p2 represents the homozygous dominant condition. By the same logic, q2 represents the homozygous recessive condition.
Let’s say that you are told that a population of acacia trees is 16 percent short (which is a, recessive) and 84 percent tall (which is A, dominant). What are the frequencies of the two alleles? Remember that it is not 0.16 and 0.84 because there are also the heterozygotes to consider!
In a problem like this, it is important to determine the value of q first because we know that all individuals with the recessive phenotype must be aa (q2). You cannot begin by calculating the value of p because it is not true that all the individuals with the dominant phenotype can be lumped into p2. Some folks displaying the dominant phenotype are heterozygous Aa (pq).
We know that q2 = 0.16, so we find q by calculating ![]() Now remember that they do not let you use a calculator. So these problems will give numbers that are fairly easy to work with. Do not despair.
Now remember that they do not let you use a calculator. So these problems will give numbers that are fairly easy to work with. Do not despair.
What about p? Since p + q is 1, and we know q = 0.40, then p must equal 1 — 0.40 or 0.600.
You may also be asked to go a step further and give the percentages of the homozygous dominant and heterozygous conditions (remember, we know that the recessive condition is 16 percent—all these individuals must be aa in order to express the recessive trait). This is simple—just plug in what you know about p and q:
2pq = (2) (0.6) (0.4) = 0.48 or 48%
p2 = (0.6) (0.6) = 0.36 or 36%
Now check your math: do the frequencies add up to 100 percent?
16 + 48 + 36 = 100
Why do we ever use the Hardy-Weinberg equation if it rarely applies to real populations? This can be an excellent tool to determine whether a population is evolving or not; if we find that the allele frequencies do not add up to one, then we need to look for the reasons for this (perhaps the population is too small and genetic drift is a factor, or perhaps one of the alleles is advantageous and is therefore being selected for and increasing in the population). Therefore, although the Hardy-Weinberg equilibrium is largely theoretical, it does have some important uses in evolutionary biology. Figure 11.7 shows an example of the Hardy-Weinberg equation in use!

Figure 11.7 The Hardy—Weinberg equilibrium. In the absence of factors that alter them, the frequencies of gametes, genotypes, and phenotypes remain constant generation after generation. (Reproduced with permission from Raven P, Johnson G, Mason K, Losos J, Duncan T; Biology, 12th ed. New York: McGraw Hill; 2020)
The Evidence for Evolution
Support for the theory of evolution can be found in varied kinds of evidence:
EVO-1
Evolution is characterized by a change in the genetic makeup of a population over time and is supported by multiple lines of evidence.
1. Homologous characters. Traits are said to be homologous if they are similar because their host organisms arose from a common ancestor (which implies that they have evolved). For example, the bone structure in bird wings is homologous in all bird species.
2. Embryology. The study of embryos reveals remarkable similarities between organisms at the earliest stages of life, although as adults (or even at birth) the species look completely different. Human embryos, for example, actually have gills for a short time during early development, hinting at our aquatic ancestry. Darwin used embryology as an important piece of evidence for the process of evolution. In 1866, the scientist Ernst Haeckel uttered the phrase, “Ontogeny recapitulates phylogeny.” Ontogeny is an individual’s development; phylogeny is a species’ evolutionary history. What Haeckel meant was that during an organism’s embryonic development, it will at some point resemble the adult form of all its ancestors before it. For example, human embryos at some point look a lot like fish embryos. The important conclusion from this is that Haeckel and others thought that embryologic similarity between developing individuals could be used to deduce phylogenetic relationships. By the end of the nineteenth century, it was clear this law rarely holds. The real development of organisms differs in several important ways from Haeckel’s schemes.
3. Vestigial characters. Most organisms carry characters that are no longer useful, although they once were. This should remind you of our short discussion about why organisms are not perfectly adapted to their environments (because the environment is constantly changing). Sometimes an environment changes so much that a trait is no longer needed, but is not deleterious enough to actually be selected against and eliminated. Darwin used vestigial characters as evidence in his original formulation of the process of evolution, listing the human appendix as an example.
Keep in mind that the kinds of evidence we’ve described are often found in the fossil record—the physical manifestation of species that have gone extinct (including things like bones as well as imprints). The most important thing to remember is that adaptations are the result of natural selection.
Phylogeny
EVO-3
Life continues to evolve within a changing environment.
Systematics is the study of evolutionary relationships and looks at the similarities and differences between species. With the similarities and differences established, you can construct phylogenetic trees and cladograms that show evolutionary relationships among lineages. While both show relationships between lineages, phylogenetic trees show the amount of change over time calibrated by fossils or a molecular clock.
Phylogenetic trees and cladograms (Figure 11.8 are used to represent evolutionary relationships among organisms as well as track traits that are either lost or gained over time. Shared characteristics will be found in multiple lineages. Shared, derived characteristics are shared only by the subset of the species and are not inherited from the most recent common ancestor.
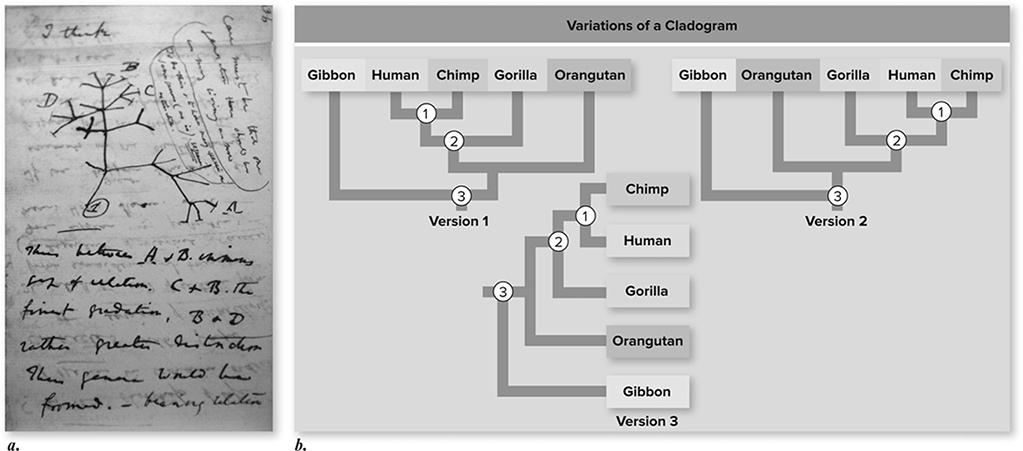
Figure 11.8 Phylogenies depict evolutionary relationships. a. A drawing from one of Darwin’s notebooks, written in 1837 as he developed his ideas that led to On the Origin of Species. Darwin viewed life as a branching process akin to a tree, with species on the twigs, and evolutionary change represented by the branching pattern displayed by a tree as it grows. b. An example of a phylogeny. Note that these three versions convey the same information despite the differences in arrangement of species and orientation. Humans and chimpanzees are more closely related to each other than they are to any other living species. This is apparent because they share a common ancestor (the node labeled 1) that was not an ancestor of other species. Similarly, humans, chimpanzees, and gorillas are more closely related to one another than any of them is to orangutans because they share a common ancestor (node 2) that was not ancestral to orangutans. Node 3 represents the common ancestor of all apes. At each node, the two descendants can be rotated without changing the meaning. For example, one difference between versions 1 and 2 is that the descendants of node 2 have been rotated so that gorilla branches to the right in version 1 and to the left in version 2. However, this does not affect the interpretation that humans and chimps are more closely related to each other than either species is to gorillas. (Reproduced with permission from Raven P, Johnson G, Mason K, Losos J, Duncan T; Biology, 12th ed. New York: McGraw Hill; 2020)
Key Ideas
The construction and/or analysis of phylogenetic trees and cladograms (Figure 11.9 is crucial on the AP exam. Keep the following in mind when constructing or analyzing them:
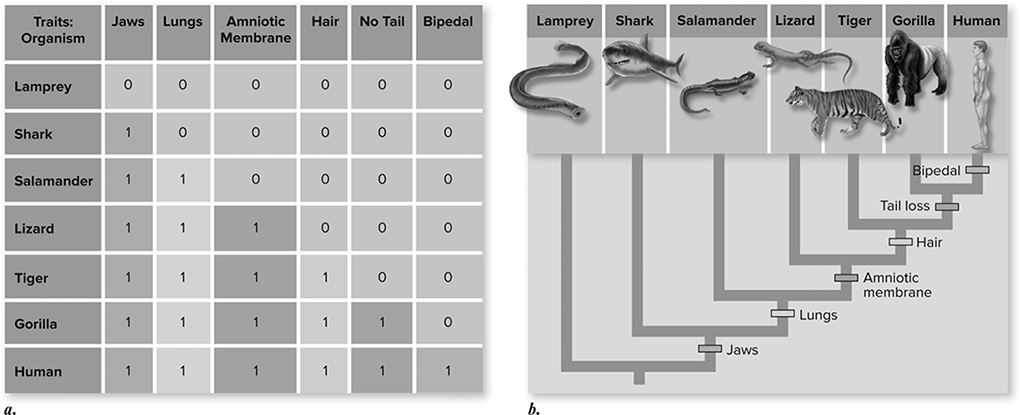
Figure 11.9 A cladogram. a. Morphological data for a group of seven vertebrates are tabulated. A “1” indicates possession of the derived character state, and a “0” indicates possession of the ancestral character state (note that the derived state for character “no tail” is the absence of a tail; for all other traits, absence of the trait is the ancestral character state). >b. A tree, or cladogram, diagrams the relationships among the organisms based on the presence of derived characters. The derived characters between the cladogram branch points are shared by all organisms above the branch points and are not present in any below them. The outgroup (in this case, the lamprey) does not possess any of the derived characters. (Reproduced with permission from Raven P, Johnson G, Mason K, Losos J, Duncan T; Biology, 12th ed. New York: McGraw Hill; 2020)
✵ Be able to read a derived characteristic chart.
✵ Nodes or branching points represent the most recent common ancestor of any two groups or lineage.
✵ Morphological similarities of living or fossil species can be used to construct phylogenetic trees or cladograms.
✵ Molecular data such as DNA or protein sequences can be used to construct phylogenetic trees or cladograms (Figure 11.10.
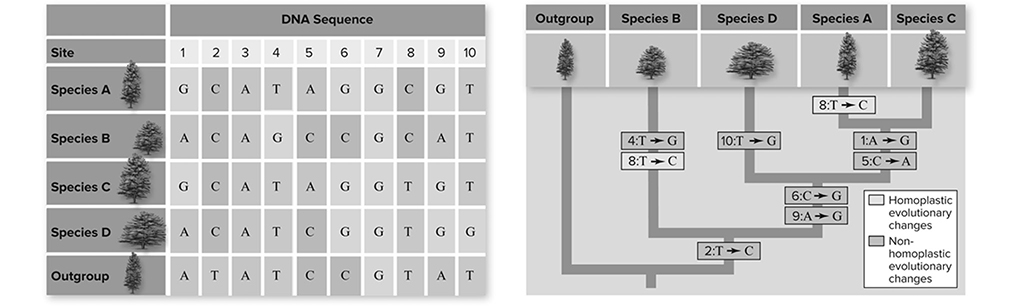
Figure 11.10 Cladistic analysis of DNA sequence data. Sequence data are analyzed just like any other data. The most parsimonious interpretation of the DNA sequence data requires nine evolutionary changes. Each of these changes is indicated on the phylogeny. Change in site 8 is homoplastic: Species A and B independently evolved from thymine to cytosine at that site. Nonhomoplastic changes are those in which all species that possess the trait derived it from the same common ancestor. Such traits are referred to as homologous. (Reproduced with permission from Raven P, Johnson G, Mason K, Losos J, Duncan T; Biology, 12th ed. New York: McGraw Hill; 2020)
✵ Phylogenetic trees and cladograms are constantly being revised and revisited as new evidence is presented.
Molecular data is the most accurate and reliable evidence for constructing phylogenetic trees or cladograms. In the following figures, a classic analysis of DNA sequence data of five different tree species is used to establish evolutionary relationships.
Macroevolution
Biologists distinguish between microevolution and macroevolution. Microevolution includes all of what we have been discussing so far in this chapter—evolution at the level of species and populations. Think of macroevolution as the big picture, which includes the study of evolution of groups of species over very long periods of time.
EVO-3
Life continues to evolve within a changing environment.
There are disagreements in the field as to the typical pattern of macroevolution. Those who believe in gradualism assert that evolutionary change is a steady, slow process, while those who think that evolution is best described by the punctuated equilibria model believe that change occurs in rapid bursts separated by large periods of stasis (no change) (see comparison in Figure 11.11). Because the fossil record is incomplete, it is very hard to test the two theories—if we find no fossils for a species over a contested period, how can we determine whether change was occurring? The debate therefore continues.

Figure 11.11 Gradualism versus punctuated equilibrium. (Reproduced with permission from Raven P, Johnson G, Mason K, Losos J, Duncan T; Biology, 12th ed. New York: McGraw Hill; 2020)
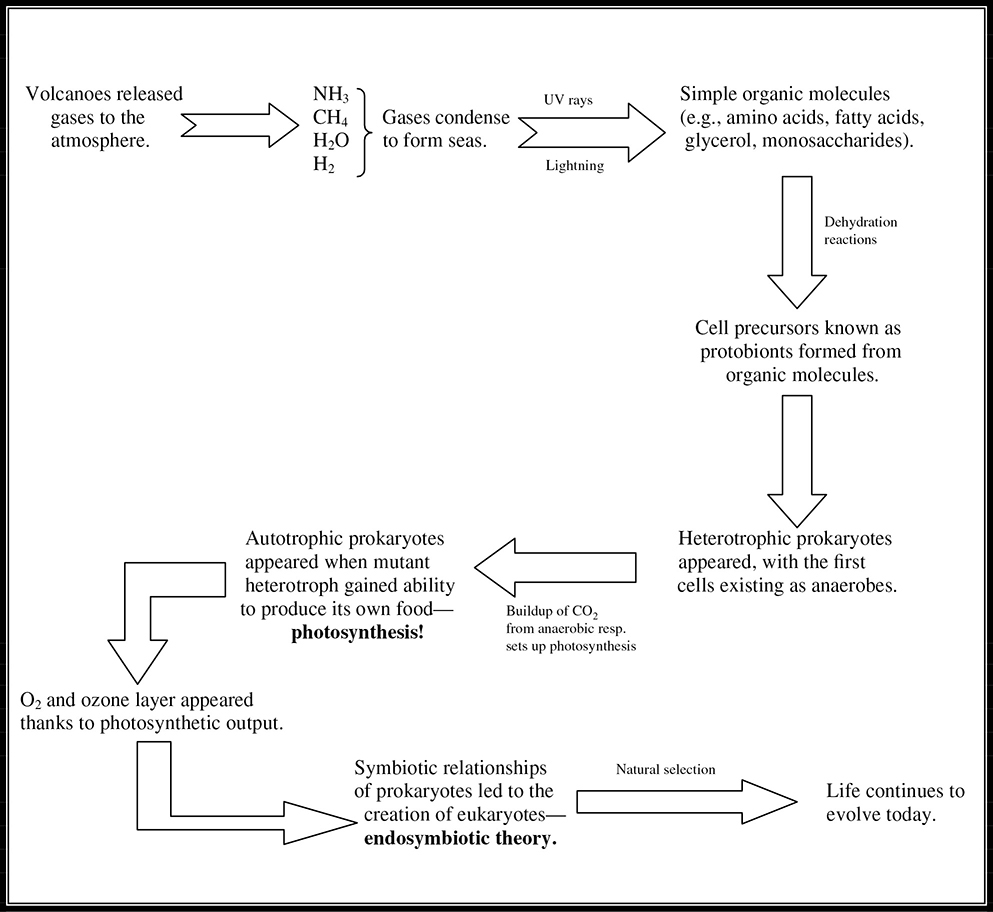
Figure 11.12 Flowchart representation of heterotroph theory.
Origins of Life on Earth
The AP Biology exam often includes questions on how life originated. It is therefore wise to learn the steps of the heterotroph theory (Figure 11.12, so named because it posits that the first organisms were heterotrophs, organisms that cannot make their own food. Geological evidence suggests.
SYI-3
Naturally occurring diversity among and between components within biological systems affects interactions with the environment.
First, though, it is important to understand that geological evidence provides support for the models of the origin of life on Earth (Figure 11.13 with Earth forming about 4.6 billion years ago with no signs of life due a hostile environment until 3.9 billion years ago. The earliest fossils date back to 3.5 billion years ago. The Miller-Urey Experiment demonstrated that several organic compounds could be formed spontaneously by simulating the conditions of Earth’s early atmosphere.
Another theory is the RNA world hypothesis, which states that RNA could have been the earliest genetic material on Earth. A simple RNA molecule could copy itself without other molecules to help, drive chemical reactions like proteins, and be able to store genetic information just like DNA.
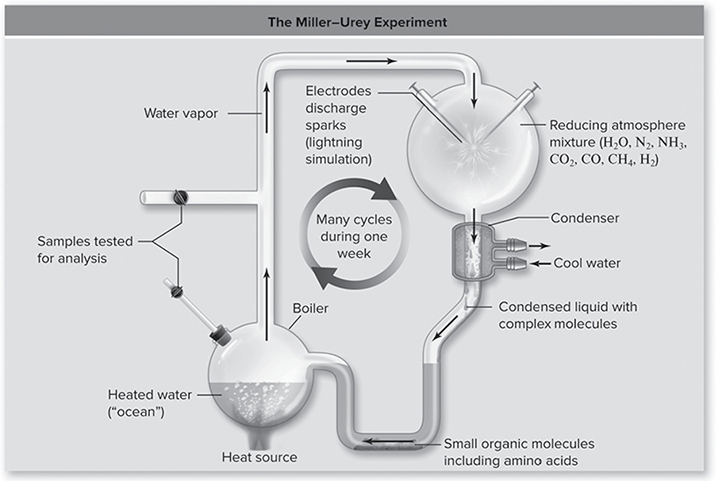
Figure 11.13 The Miller—Urey experiment. The apparatus consisted of a closed tube connecting two chambers. The upper chamber contained a mixture of gases thought to resemble the primitive Earth’s atmosphere. Electrodes discharged sparks through this mixture, simulating lightning. Condensers then cooled the gases, causing water droplets to form, which passed into the second heated chamber, the “ocean.” Any complex molecules formed in the atmosphere chamber would be dissolved in these droplets and carried to the ocean chamber, from which samples were withdrawn for analysis. (Reproduced with permission from Raven P, Johnson G, Mason K, Losos J, Duncan T; Biology, 12th ed. New York: McGraw Hill; 2020)
![]() Review Questions
Review Questions
1. Which of the following is an evolutionary process not based on random factors?
A. Genetic drift
B. Natural selection
C. Mutation
D. Gene flow
2. Which of the following is not a sexually selected trait?
A. Fruit fly wings
B. A male baboon’s canine teeth
C. Peacock tail feathers
D. Male/female dimorphism in body size in many species
3. An adaptation
A. can be shaped by genetic drift.
B. cannot be altered.
C. evolves because it specifically improves an individual’s mating success.
D. affects the fitness of an organism if it is altered.
4. Which of the following is not a requirement for natural selection to occur?
A. Variation between individuals
B. Heritability of the trait being selected
C. Sexual reproduction
D. Differences in reproductive success among individuals
5. Why can Hardy-Weinberg equilibrium occur only in large populations?
A. Large populations are likely to have more variable environments.
B. More individuals means less chance for natural selection to occur.
C. Genetic drift is a much stronger force in small versus large populations.
D. Large populations make random mating virtually impossible.
6. A population of frogs consists of 9 percent with speckles (the recessive condition) and 91 percent without speckles. What are the frequencies of the p and q alleles if this population is in Hardy- Weinberg equilibrium?
A. p = 0.49, q = 0.51
B. p = 0.60, q = 0.40
C. p = 0.70, q = 0.30
D. p = 0.49, q = 0.30
7. Frequency-dependent selection is
A. particularly important during speciation.
B. one way in which multiple alleles are preserved in a population.
C. possible only when there are two alleles.
D. most common in bacteria.
8. All of the following provide evidence for evolution except
A. vestigial characters.
B. Darwin’s finches.
C. homologous characters.
D. mutations.
9. Why do we assume that oxygen was not present in the original atmosphere?
A. The presence of O2 would have resulted in the evolution of too many species too fast.
B. Oxygen would have slowed down the rate of evolution.
C. We know the ozone layer, which is formed by oxygen, has not been around that long.
D. Inorganic molecules could not have formed in the presence of oxygen.
10. All of these are examples of random evolutionary processes except
A. an earthquake divides a single elk species into two populations, forcing them to no longer interbreed.
B. a mutation in a flower plant results in a new variety.
C. an especially long winter causes a group of migrating birds to shift their home range.
D. a mutation results in a population of trees that spread their seeds more widely than their peers, causing their population to grow.
![]() Answers and Explanations
Answers and Explanations
1. B—Natural selection is the selective increase in certain alleles because they confer an advantage to their host organism. All other factors are random with respect to the alleles (a “bottleneck” is a type of genetic drift where a population is drastically reduced in size).
2. A—All fruit flies need to fly not only to find mates but also to survive. All the other characters listed are sexually selected, meaning that they have evolved because they confer specific advantages in mating (and not survival).
3. D—Adaptations are defined as traits that affect fitness if they are altered. Although adaptations may have evolved to increase mating success (answer C), they are not always intended for that function (e.g., they may have remained because they increase survival).
4. C—Natural selection can occur in asexually reproducing organisms, as long as the other three necessary conditions are met.
5. C—Genetic drift is change in allele frequencies as a result of random factors (e.g., natural disasters or environmental change). In small populations, genetic drift is a much more powerful force because each individual represents a greater percentage of the population’s total genes than that person would in a much larger population. Think of it this way—if you have a population of 10 cheetahs, and 3 die, you have lost 30 percent of the genes in that pool. If you have a population of 100 cheetahs, and 3 die, you have lost only 3 percent. Since Hardy-Weinberg equilibrium depends on no genetic drift, it is much more likely to occur in very large populations.
6. C—Remember that p and q must add up to 1 for a population to be in Hardy-Weinberg equilibrium (this eliminates answer D). Calculate q first by taking the square root of 0.09, which is 0.30. Then simply subtract 0.30 from 1 to get p = 0.70.
7. B—Frequency-dependent selection is one process by which multiple alleles are preserved in a population. For traits that are selected for or against on the basis of frequency, an allele becomes more advantageous when it is rare, and therefore increases. In this way, it is impossible for the allele to become extinct (because as soon as it gets that low, it increases again). When it gets too high, the other allele is low, and that one then increases. Frequency-dependent selection often exhibits itself in this kind of seesaw effect.
8. D—Mutations in and of themselves are not evidence for evolution, although they are necessary if evolution is going to occur.
9. D—Inorganic molecules could not have formed in the presence of oxygen because oxygen would have taken the place of other elements in every chemical reaction (because it is such a highly reactive element).
10. D—This is the only answer that shows evidence of natural selection, which is the nonrandom process by which evolution occurs. The two elk species splitting (answer A) is an example of allopatric speciation caused by a random factor (a geologic event). A mutation is also a random event (answer B); for example, if we had said that the new variety became the dominant allele in a population because it had an advantage over other variants, then that would be natural selection. A home range shift (answer C) is not evolution, but rather a behavioral change within an organism’s lifetime.
![]() Rapid Review
Rapid Review
There are four modes of evolution:
1. Genetic drift: change in allele frequencies because of chance events (in small populations).
2. Gene flow: change in allele frequencies as genes move from one population to another.
3. Mutation: change in allele frequencies due to a random genetic change in an allele.
4. Natural selection: process by which characters or traits are maintained or eliminated in a population based on their contribution to the differential survival and reproductive success of their “host” organisms.
There are three requirements for natural selection to occur:
1. Variation: differences must exist between individuals.
2. Heritability: the traits to be selected for must be able to be passed along to offspring. Traits that are not inherited cannot be selected against.
3. Differential reproductive success: there must be variation among parents in how many offspring they produce as a result of the different traits that the parents have.
Adaptation is a trait that, if altered, affects the fitness of an organism; includes physical or intangible traits.
Selection types are as follows:
1. Directional: members at one end of a spectrum are selected against, and population shifts toward that end.
2. Stabilizing: selection for the mean of a population; reduces variation in a population.
3. Disruptive (diversifying): selects for the two extremes of a population; selects against the middle.
4. Sexual: certain characters are selected for because they aid in mate acquisition.
5. Artificial: human intervention in the form of selective breeding (cattle).
Sources of variation within populations include the following:
1. Mutation: random changes in DNA can introduce new alleles into a population.
2. Sexual reproduction: crossover, independent assortment, random gamete combination.
3. Balanced polymorphism: the maintenance of two or more phenotypic variants.
Speciation is the process by which new species evolve:
1. Allopatric speciation: interbreeding stops because some physical barrier splits the population into two. If two populations evolve separately and change so they cannot interbreed, speciation has occurred.
2. Sympatric speciation: interbreeding stops even though no physical barrier prevents it.
✵ Polyploidy: condition in which individual has a higher-than-normal number of chromosome sets. Polyploidic individuals cannot reproduce with nonpolyploidics.
✵ Balanced polymorphism: two phenotypic variants become so different that the two groups stop interbreeding.
Other terms to remember include the following:
Adaptive radiation: rapid series of speciation events that occur when one or more ancestral species invades a new environment.
Hardy-Weinberg equilibrium: p + q = 1, p2 + 2pq + q2 = 1. Evolution is not occurring. The rules for this are no mutations, no gene flow, no genetic drift, no natural selection, and random mating.
Homologous character: traits similar between organisms that arose from a common ancestor.
Vestigial character: character contained by organism that is no longer functionally useful (appendix).
Gradualism: evolutionary change is a slow and steady process.
Punctuated equilibria: evolutionary change occurs in rapid bursts separated by large periods of no change.
Heterotroph theory: theory that describes how life evolved from original heterotrophs.
Convergent character: traits similar to two or more organisms that do not share a common ancestor; parallel evolution.
Convergent evolution: two unrelated species evolve in a way that makes them more similar.
Divergent evolution: two related species evolve in a way that makes them less similar.