Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Plant Structure
Structure of Woody Plants

Chapter Opener Image: These are relatively small trunks of western red cedars (Thuja plicata), they become more massive with age. Each year, the trunk, branches, and roots add a new layer of wood and bark, making them stronger and more conductive to water, minerals, and sugars. With such strength, the trees grow tall enough not only to put their leaves above those of shorter plants, they also put their reproductive cones higher, such that there is a better chance wind will carry pollen from each tree to many others.
OUTLINE
✵ Concepts
✵ Vascular Cambium
- Initiation of the Vascular Cambium
- Fusiform Initials
- Ray Initials
- Arrangement of Cambial Cells
✵ Secondary Xylem
- Types of Wood Cells
- Growth Rings
- Heartwood and Sapwood
- Reaction Wood
✵ Secondary Phloem
✵ Outer Bark
- Cork and the Cork Cambium
- Lenticels and Oxygen Diffusion
- Initiation of Cork Cambia
✵ Secondary Growth in Roots
✵ Anomalous Forms of Growth
- Anomalous Secondary Growth
- Unusual Primary Growth
Box 8-1 Botany and Beyond: Wood in Three Dimensions
Box 8-2 Plants Do Things Differently: Having Multiple Bodies in One Lifetime
Box 8-3 Plants and People: Dendrochronology—Tree Ring Analysis
Box 8-4 Plants Do Things Differently: Thinking about the Growth of Wood
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Explain the advantages and disadvantages of secondary growth.
✵ Define the vascular cambium, fusiform initials, and ray initials.
✵ Describe the arrangement of cambial cells.
✵ Discuss the axial system of wood.
✵ Identify the portions of growth rings.
✵ Define heartwood, sapwood, and reaction wood.
✵ Compare secondary phloem to secondary xylem.
✵ Contrast cork cambium and vascular cambium.
✵ Explain the function of lenticels.
✵ Discuss secondary root growth.
✵ Name three examples of anomalous secondary growth.
✵ Give an example of establishment growth.
 Did You Know?
Did You Know?
✵ Woody plants are usually easy to recognize: If you think a tree or shrub is woody, you are probably correct.
✵ Only a few plants will fool you: Palms and bamboos have no wood, and many plants that appear to be herbs (they die after a few months and decompose during winter) have a small amount of delicate wood.
✵ Most woody plants add a new layer of wood (a growth ring) around their trunk, branches, and roots every year.
✵ There is no theoretical limit to how long a plant can live and how wide it can become; a cypress in Oaxaca, Mexico has a diameter of 11.6 m (38 feet).
![]() Concepts
Concepts
Growth by means of apical meristems has been previously described. From the meristems are derived sets of tissues: epidermis, cortex, vascular bundles, pith, and leaves. These primary tissues together constitute the primary plant body. In plants known technically as herbs, this is the only body that ever develops, but in woody species, additional tissues are produced in the stem and root from other meristems—the vascular cambium and the cork cambium. The new tissues themselves are wood (secondary xylem) and bark (secondary phloem and cork); they are secondary tissues, and they constitute the plant’s secondary body. Examples of woody plants are abundant: Trees such as sycamores, chestnuts, pines, and firs are woody, as are shrubs such as roses, oleanders, and azaleas.
The ability to undergo secondary growth and produce a woody body has many important consequences. In an herb, after a portion of stem or root is mature, its conducting capacity is set. All provascular cells have differentiated into either primary xylem or primary phloem. This capacity is correlated with the needs of leaves and roots. If the plant produces so many leaves that they lose water faster than the stem xylem can conduct, some or all the leaves die of water loss. Similarly, it would not be selectively advantageous for the plant to have so many leaves that they could produce sugar faster than the phloem could conduct it to roots, flower buds, or developing fruits.
Many herbs live for several years, however. How do they respond to the bottleneck of the limited conducting capacity of the first-year stem? In some, the first year’s leaves die during winter, and in the second year, the plant produces only as many leaves as it had during the first year. In other species, adventitious roots are produced that supply conducting capacity directly to the new section of stem being formed, thus bypassing older portions of the stem. Most of these plants must remain low enough for adventitious roots to reach the soil. These are often rhizomatous, such as irises, bamboo, and ferns.
Woody plants not only become taller through growth by their apical meristems but also become wider by accumulation of wood and bark. Because wood and bark contain conducting tissues, their accumulation gives plants a greater capacity to move water and minerals upward and carbohydrates downward. The number of leaves and roots that the plant can support increases, as does the photosynthetic capacity.
For example, consider a tree with a trunk radius of 10.5 cm (FIGURE 8-1); imagine that the plant produced a layer of wood 0.5 cm thick in the previous year. As a consequence, the tree has 32.2 cm2 of new wood that conducts water from roots to leaves. Assume that each leaf loses water at a rate equal to the conducting capacity of 0.1 cm2 of wood; this plant can conduct enough water through its trunk to support 322 leaves. If the tree produces another 0.5 cm of wood this year, the new ring of wood will have a cross-sectional area of 33.8 cm2. It is larger than the previous ring of wood and can support conduction to a greater number of leaves—338. Even if a ring of wood could conduct for only 1 year, the plant could still produce a greater number of leaves every year, and thus, its annual photosynthetic capacity would always increase. The consequence of this ever-increasing capacity is that annual production of seeds and defensive chemicals also increases.
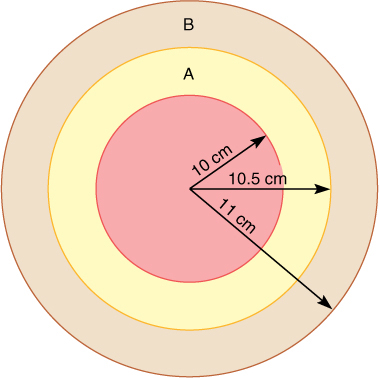
FIGURE 8-1 The cross-sectional area of a ring of wood is given by the formula p times the square of the outer radius minus the square of the inner radius. If each annual ring is 0.5 cm wide, when the wood has a radius of 10.5 cm, its newest ring has a cross-sectional area of 32.2 cm2. Next year’s ring will be larger, 33.8 cm2, an increase of (33.8 — 32.2)/32.2 = 1.6/32.2 3 100% = 5% (not drawn to scale).
Only those seeds that germinate in a suitable site are able to grow into adults and reproduce. After a seed of a woody, perennial plant germinates and becomes established, it occupies its favorable site year after year. Other seeds may not be able to germinate because they do not encounter suitable sites. In springtime, all sites that had been occupied by herbs are vacant and available to the seeds of both annual herbs and woody perennials, but virtually all sites occupied by last year’s perennials are still occupied by them. Many pines, oaks, and other long-lived trees hold on to the same piece of Earth for as long as 600 years and a few for up to 3,000 years; one particular tree has lived for 5,000 years. For all of that time the trees produce seeds of their own and, by their very presence, prevent the seeds of their competitors from growing at that site. Annual herbs give up their sites when they die; their seeds must compete for new sites every year.
Secondary growth also has disadvantages: A 5,000-year-old plant is 10,000 times older than an herb that germinates in April, lives 6 months, then sets seed and dies by September. It has had to battle insects, fungi, and environmental harshness 10,000 times longer, and it is a bigger, more easily discovered target for pathogens. Perennials have a greater need for defenses, both chemical and structural, than annual herbs have, and they must use a portion of their energy and nutrient resources for winterizing their bodies if they live in temperate climates. It is also expensive metabolically to construct wood and bark. The fact that wood burns so readily shows that it is energy rich. If no secondary growth occurred, this energy could be used immediately for reproduction. In fact, most woody plants do not reproduce until they are several years old; if they are killed by disease or environmental stress before they reproduce, all growth and development have been for nothing.
It must be difficult for secondary growth to arise by evolution; it has originated only three times in the 420 million years that vascular plants have been in existence, and two of those evolutionary lines later became extinct. All woody trees and shrubs alive today have descended from just one group of ancestral woody plants that arose about 370 million years ago. This group has been very successful, evolving into many species and dominating almost all regions of Earth; they include all seed plants. Within the flowering plants, herbaceousness is a new phenomenon; all early angiosperms were woody perennials, but many plants have evolved to be herbs, foregoing the woody life style. Currently, true secondary growth occurs in many eudicots, most basal angiosperms, and all gymnosperms, but not in any ferns or monocots (TABLE 8-1).
As you study this, remember that a woody plant is a combination of primary and secondary tissues: The tips of stems and roots, as well as the leaves, flowers, and fruits, are herbaceous and primary; only as portions of stems and roots become older do they begin to undergo secondary growth and become woody.
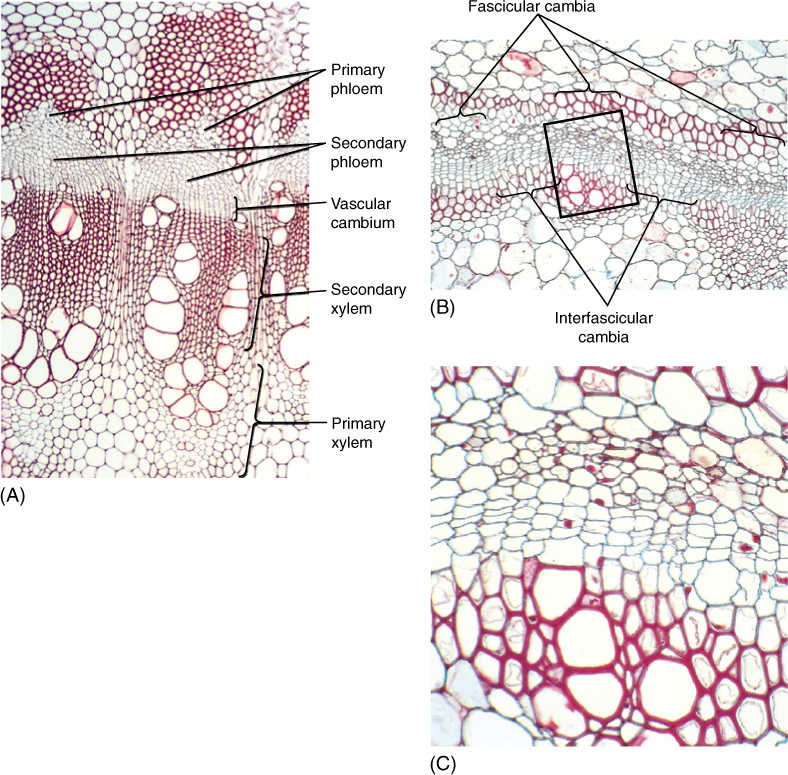
FIGURE 8-2 (A) The cambium has recently formed in this Bocconia stem. The fascicular cambia are rather obvious, but the interfascicular cambia are difficult to see. They have produced more secondary xylem than secondary phloem (×180). (B) and (C) This cambium has only recently formed; it has produced two or three layers of secondary xylem and one or two of secondary phloem. Three groups of primary xylem are visible, and thus, you can tell where fascicular and interfascicular cambia arose. The boxed area in (B) is magnified in (C); (B) ×50; (C) ×250.
TABLE 8-1 Presence of Secondary Growth

![]() Vascular Cambium
Vascular Cambium
Initiation of the Vascular Cambium
The vascular cambium (plural, cambia) is one of the meristems that produce the secondary plant body (FIGURE 8-2). In an herbaceous species, the cells located between the metaxylem and metaphloem of a vascular bundle ultimately stop dividing and differentiate into conducting tissues. But in a woody species, the cells located in this position never undergo cell cycle arrest; they continue to divide instead of maturing, and they constitute the fascicular cambium (FIGURE 8-3A; “fascicle” is an old term for bundle). In addition, some mature parenchyma cells between vascular bundles come out of cell cycle arrest and resume mitosis, forming an interfascicular cambium that connects on each side with the fascicular cambia. After this happens, the vascular cambium is a complete cylinder. The terms “fascicular” and “interfascicular” are used only while the cambium is young; after 2 or 3 years, the two regions are usually indistinguishable, and then only the term “vascular cambium” is used.
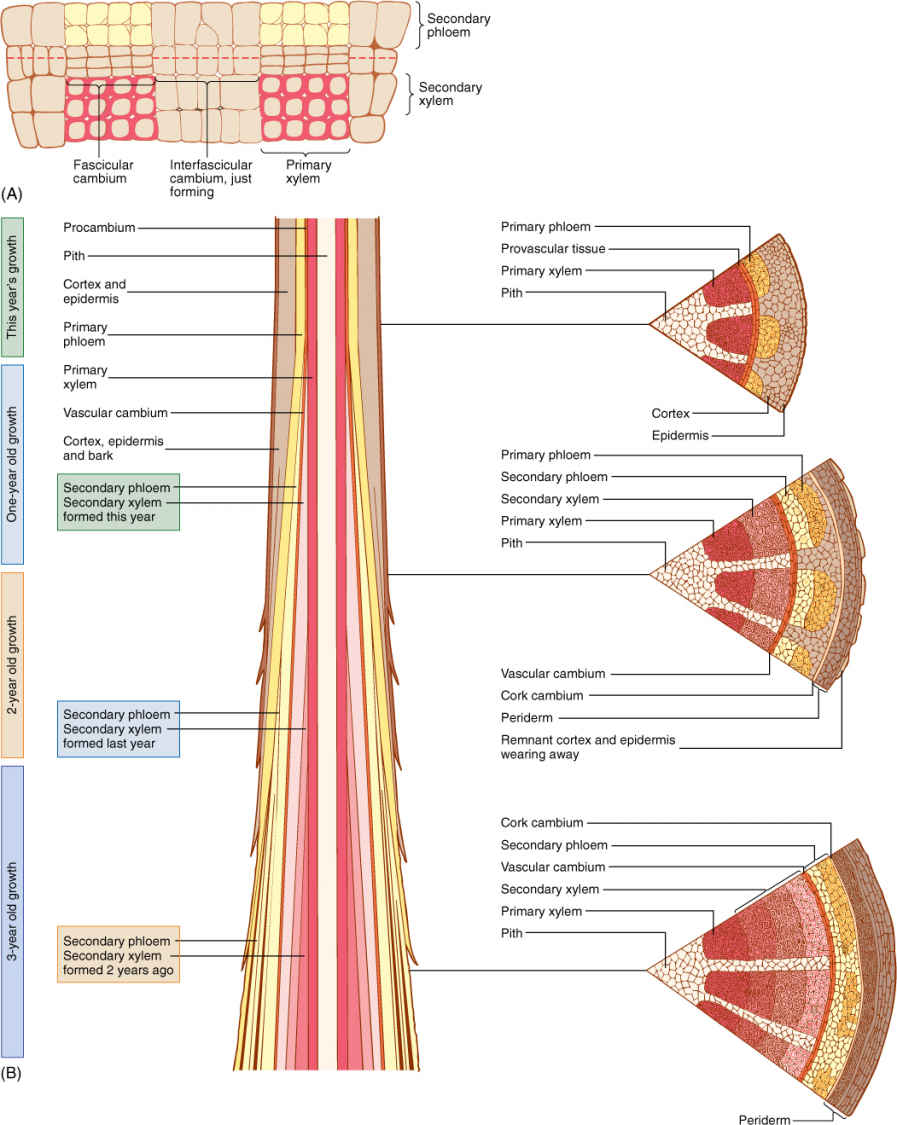
FIGURE 8-3 (A) Two vascular bundles and the parenchyma located between them. Some parenchyma cells have begun renewed cell division and constitute an interfascicular cambium. This zone of renewed mitotic activity is located between two fascicular cambia, so one full cambial zone will result. (B) The vascular cambium usually does not form until after a portion of shoot or root is several weeks or even many months old; so branch tips have only primary growth. After a region is old enough, the vascular cambium forms, and secondary xylem begins to accumulate to the inside of the cambium; secondary phloem accumulates to its exterior. At first, vascular bundles are still recognizable. After several years of activity, considerable amounts of secondary xylem and phloem accumulate.
Vascular cambia must be extended each year. The tips of roots and stems initially contain only primary tissues; however, at some time after metaxylem and metaphloem have matured, a vascular cambium arises, and that portion of the root or stem then contains both primary and secondary tissues (FIGURE 8-3B). During the next growing season, the apical meristem extends the axis beyond this point; a new segment of vascular cambium forms within it and joins at its base to the top of the vascular cambium formed in the previous season. The vascular cambium within a tree consists of segments of distinct ages, those near the ground being oldest and those closer to the tips of the axes being younger.
Very rarely, a vascular cambium forms in leaves that stay on a tree for many years, but just a tiny amount of secondary tissues, usually only secondary phloem without any secondary xylem, is formed in the midrib; the other veins contain only primary tissues. Vascular cambia never occur in flowers, fruits, or seeds.
Although a vascular cambium shares many features with an apical meristem, it is unique in certain aspects. It is a rather simple meristem in that it has only two types of cells, fusiform initials and ray initials (FIGURES 8-4 and 8-5).
Fusiform Initials
Fusiform initials are long, tapered cells; typical lengths for fusiform initials are 140 to 462 mm in dicots and 700 to 8,700 mm (almost 1 cm) in conifers. When a fusiform initial undergoes longitudinal cell division with a wall parallel to the circumference of the cambium (a periclinal wall), it produces two elongate cells (FIGURE 8-6). One continues to be a fusiform initial; the other differentiates into a cell of secondary xylem or secondary phloem. If the outer daughter cell remains a cambium cell, the inner cell develops into secondary xylem. But if the inner one continues as cambium, the outer cell differentiates and matures into secondary phloem. This orientation is constant: Wood never forms to the exterior of the vascular cambium, and bark never forms on the interior side. Regardless of which cell differentiates, one always remains as cambium. It is not known what factors determine which cell remains cambial and which differentiates, but within any year, both xylem and phloem are produced, almost always much more xylem than phloem.
Cambial cells produce narrow daughter cells, all of which enlarge during differentiation. Daughter cells located on the inner side, which mature into secondary xylem, increase greatly in diameter, causing the cambial cells to be pushed outward (FIGURES 8-7 and 8-8). Because the cambium is a cylinder, such outward movement results in a cylinder of larger circumference. Vascular cambium cells must occasionally divide longitudinally by anticlinal walls (perpendicular to the cambium’s surface), thereby increasing the number of cambial cells (Figure 8-6F). Without anticlinal divisions, cambial cells would be stretched wider circumferentially and finally could not function (FIGURE 8-9).
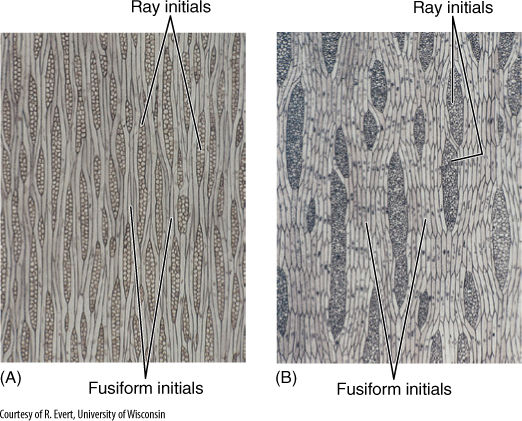
FIGURE 8-4 Tangential sections through a nonstoried vascular cambium of apple (Malus sylvestris) (A) and a storied vascular cambium of black locust (Robinia pseudoacacia) (B). The nonstoried fusiform initials are extremely long, and the ends are not aligned. In the storied cambium, fusiform initials are much shorter and occur in horizontal rows.
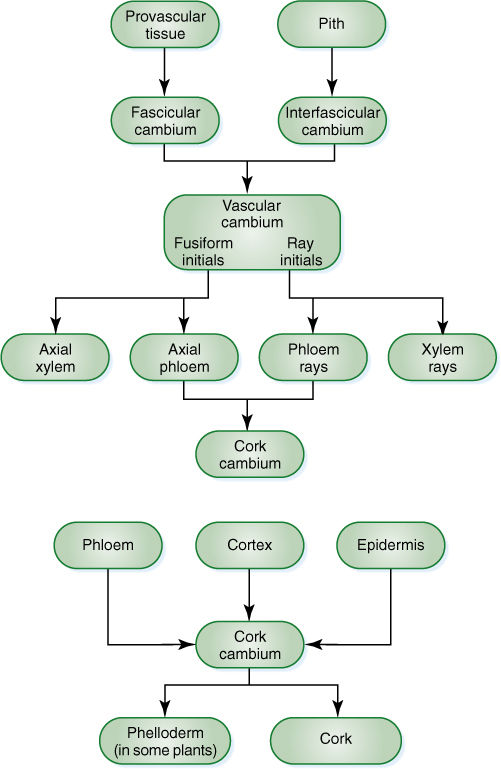
FIGURE 8-5 Cell lineages during secondary growth.
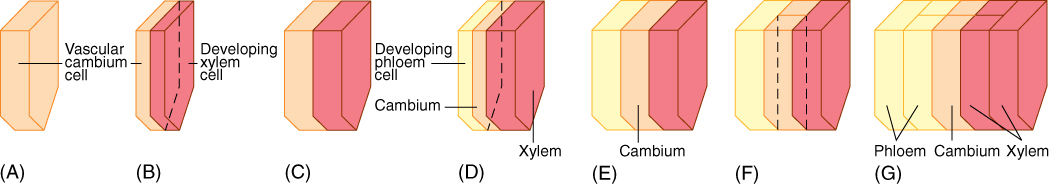
FIGURE 8-6 (A) The lower half of a fusiform initial before division (to simplify the diagram, the top half has not been drawn in). (B) Division by a periclinal wall results in two thin cells; the outer one remains a fusiform initial and the inner cell develops into secondary xylem. (C) Both cells enlarge to the size of the original cell. (D) The fusiform initial divides again; this time the outer cell matures as secondary phloem, whereas the inner one remains a fusiform initial. (E) The cells grow back to the original size. (F) The fusiform initial divides by an anticlinal wall, resulting in two fusiform initials. (G) Two division cycles after the radial division in (F), a new row of cells is present in the secondary xylem and phloem, produced by the new cambial cell formed in (F).
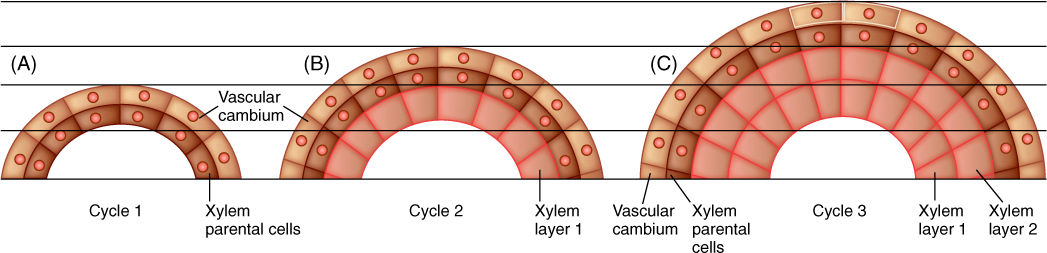
FIGURE 8-7 (A) After vascular cambium cells divide, progeny cells on the interior side become xylem parental cells. (B) As all cells expand to their mature size, the cambial cells are pushed outward. Cambial cells divide again, depositing a new layer of xylem parental cells exterior to the xylem that just formed (xylem layer 1). (C) The new xylem parental cells expand, pushing the vascular cambium farther outward. Cambial cells divide again. Each xylem cell is formed in place; they are not pushed inward.
Like apical meristem cells, fusiform initials have thin primary walls, and plastids are present as proplastids. After nuclear division, a phragmoplast forms and elongates toward the ends of the cell. The phragmoplast grows about 50 to 100 mm per hour, and cell division may take as long as 10 days in species with long fusiform initials, whereas the cell cycle may be as short as 19 hours in apical meristem cells of the same plant.
Ray Initials
Ray initials are similar to fusiform initials except that they are short and more or less cuboidal. They too undergo periclinal cell divisions, with one of the daughters remaining a cambial ray initial and the other differentiating into either xylem parenchyma if it is the inner cell, or phloem parenchyma if it is the outer cell. One of the most significant differences between fusiform and ray initials is that the elongate fusiform initials produce the elongate cells of wood (tracheids, vessel elements, and fibers) and phloem (sieve cells, sieve tube members, companion cells, and fibers). Ray initials produce short cells, mostly just storage parenchyma and, in gymnosperms, albuminous cells.
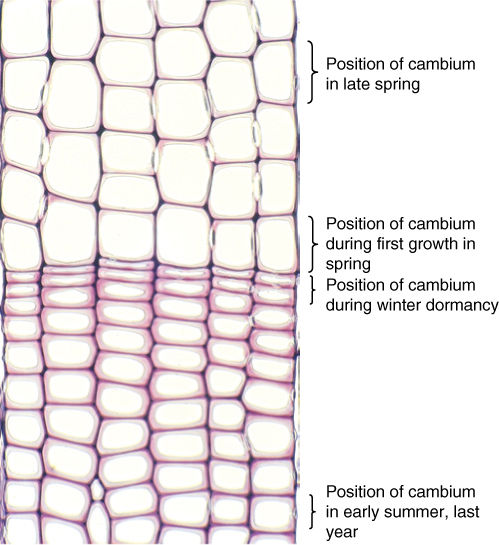
FIGURE 8-8 Six rows of wood cells are visible in this transverse section of a pine trunk. All cells of each row were produced by a single vascular cambium cell. Thus, all of the wood visible in this micrograph was produced by just six cambium cells. The cells at the bottom of the picture were produced when the cambium cells were located at that site; similarly, the cells in the middle of the picture were produced when the cambium existed there, having been pushed outward by the production of the cells at the bottom of the picture (3150).
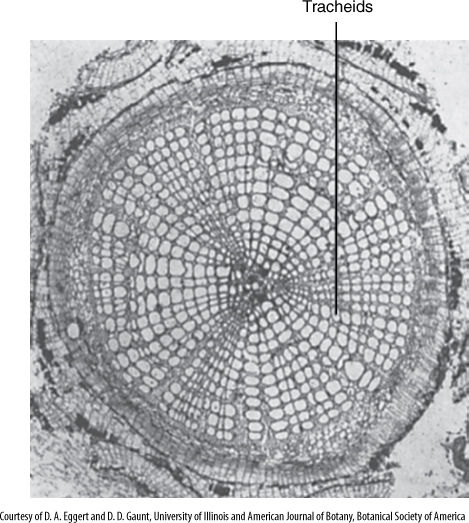
FIGURE 8-9 Many extinct arthrophytes were tree-like, having a wood of large tracheids, but their fusiform initials could not undergo longitudinal radial divisions, so no new fusiform initials could be formed and secondary growth was limited (×100).
Arrangement of Cambial Cells
Ray and fusiform initials are organized in specific patterns. Ray initials are typically grouped together in short vertical rows only one cell wide (uniseriate), two cells wide (biseriate), or many cells wide (multiseriate; see Figure 8-4). Fusiform initials may occur in regular horizontal rows (a storied cambium) or irregularly, without any horizontal pattern (a nonstoried cambium). Storied cambia have evolved more recently than nonstoried cambia and occur in only a few advanced eudicot species, for example, redbud and persimmon. The fusiform initials of storied cambia tend to be short, less than 200 mm long. The selective advantage of storied tissues is unknown.
Typically, a vascular cambium never has large regions of just fusiform initials or just ray initials. If anticlinal divisions result in many fusiform initials side by side, a central one may undergo transverse divisions and be transformed into a set of ray initials (FIGURE 8-10). Likewise, if a group of ray initials becomes unusually broad, one or several central ones may elongate and be converted to a fusiform initial. The ratio of fusiform initials to ray initials within a species is quite constant and apparently under precise developmental control.
To summarize, a single tree or shrub has only a single vascular cambium, in the form of a cylinder, one cell thick, extending upward in trunks, outward into branches almost to the very tips of each twig, and downward in roots almost to their tips. Each section of vascular cambium forms as a narrow cylinder but is pushed outward by its own production of wood cells on its inner surface.

FIGURE 8-10 A fusiform initial can divide transversely and become a row of ray initials, after which all its derivatives differentiate as ray cells.
![]() Secondary Xylem
Secondary Xylem
Types of Wood Cells
All cells formed to the interior of the vascular cambium develop into secondary xylem, known as wood. Secondary xylem contains all of the types of cells that occur in primary xylem but no new ones. Wood may contain tracheids, vessel elements, fibers, sclereids, and parenchyma. The only real differences between primary and secondary xylem are the origin and arrangement of cells. The arrangement of secondary xylem cells reflects that of the fusiform and ray initials: An axial system is derived from the fusiform initials, and a radial system develops from the ray initials (FIGURE 8-11; Box 8-1, Botany and Beyond: Wood in Three Dimensions).
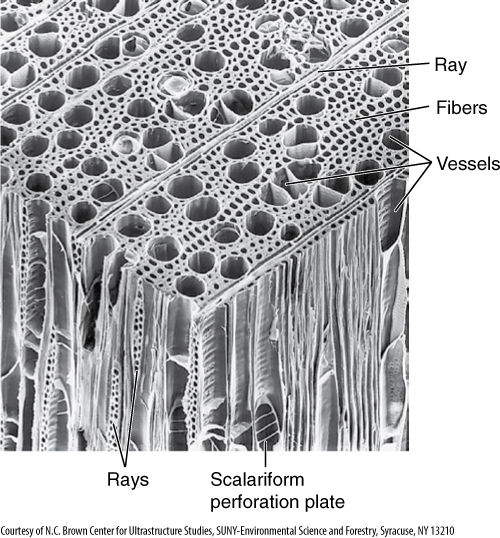
FIGURE 8-11 Scanning electron micrograph of a cube of wood from a tulip tree (Liriodendron tulipifera). Examine the relationship between the various components, especially the large vessels and the ray parenchyma (×110).
Botany and Beyond
BOX 8-1 Wood in Three Dimensions
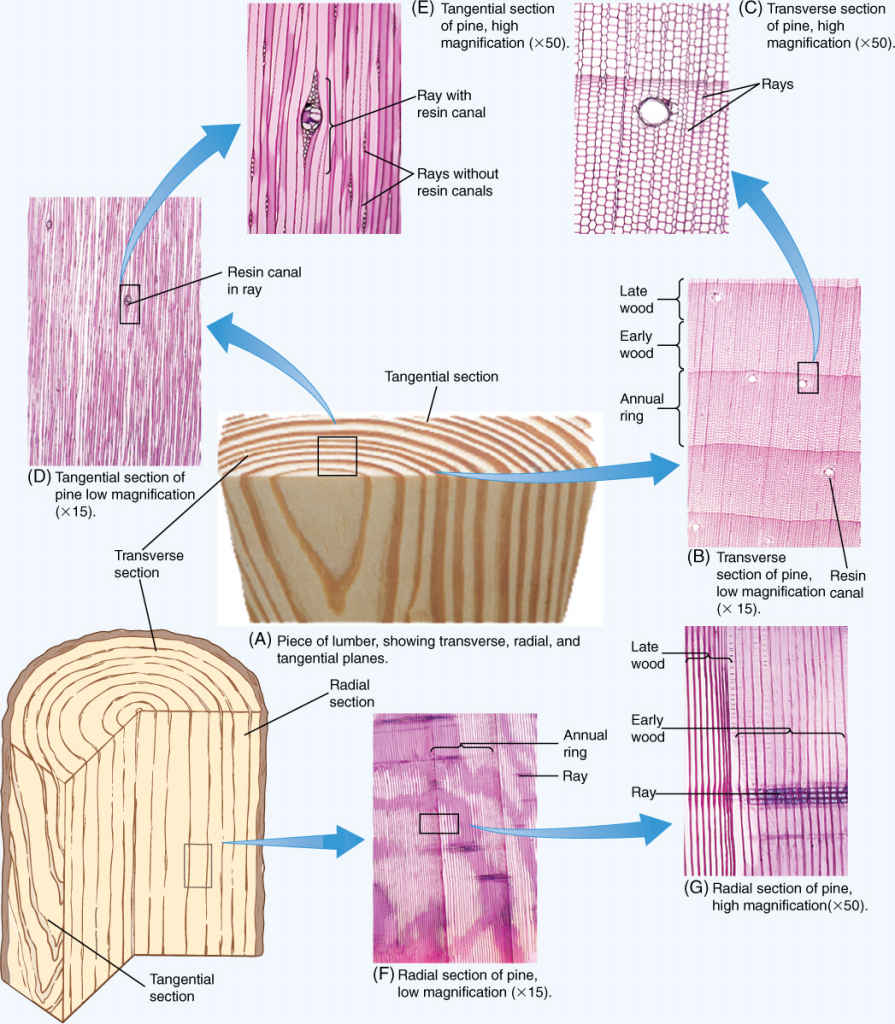
It is important to understand wood in all three dimensions, but that is not particularly easy. As you study these illustrations, think about how the cells are formed by the vascular cambium. Also try to picture how the various cells touch each other in three dimensions, which conduct water, which store water and nutrients, and so on. Pine wood is simpler because its axial system contains only tracheids, so you need to think mostly about tracheids and rays. Oak wood is more complex, with vessels, fibers, and axial parenchyma all touching the rays at some point. For some serious practice of your skill at visualizing three-dimensional objects, imagine a vascular cambium existing on one tangential face. How are the new cells formed each year?
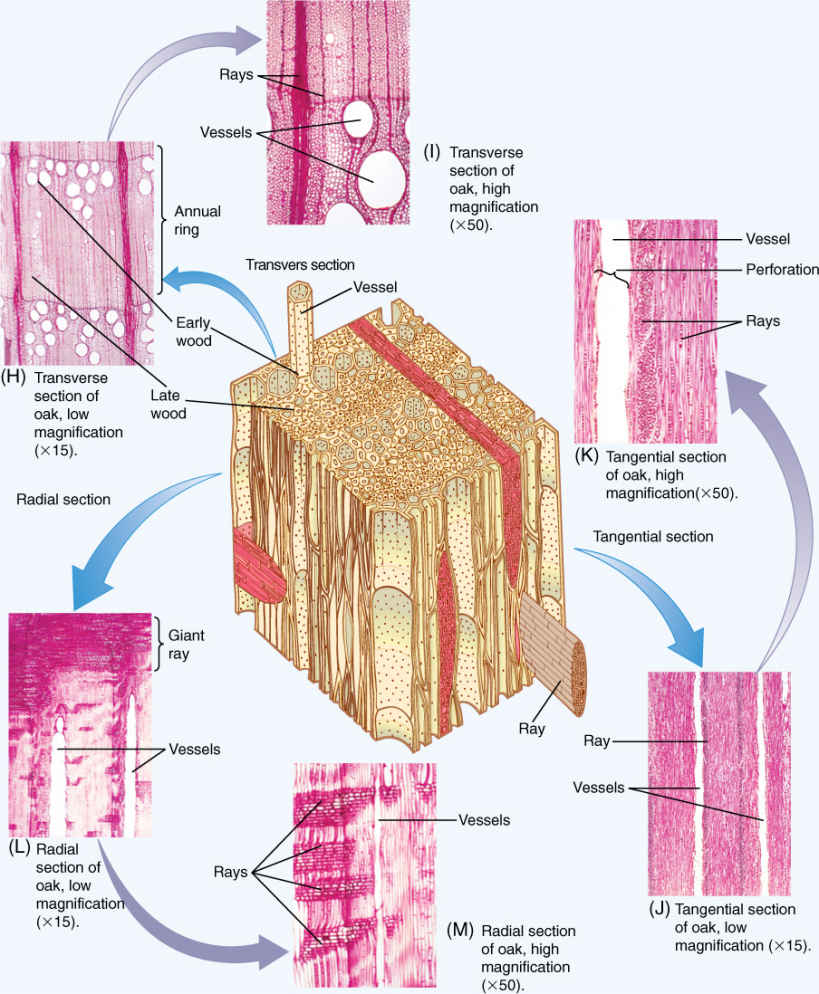
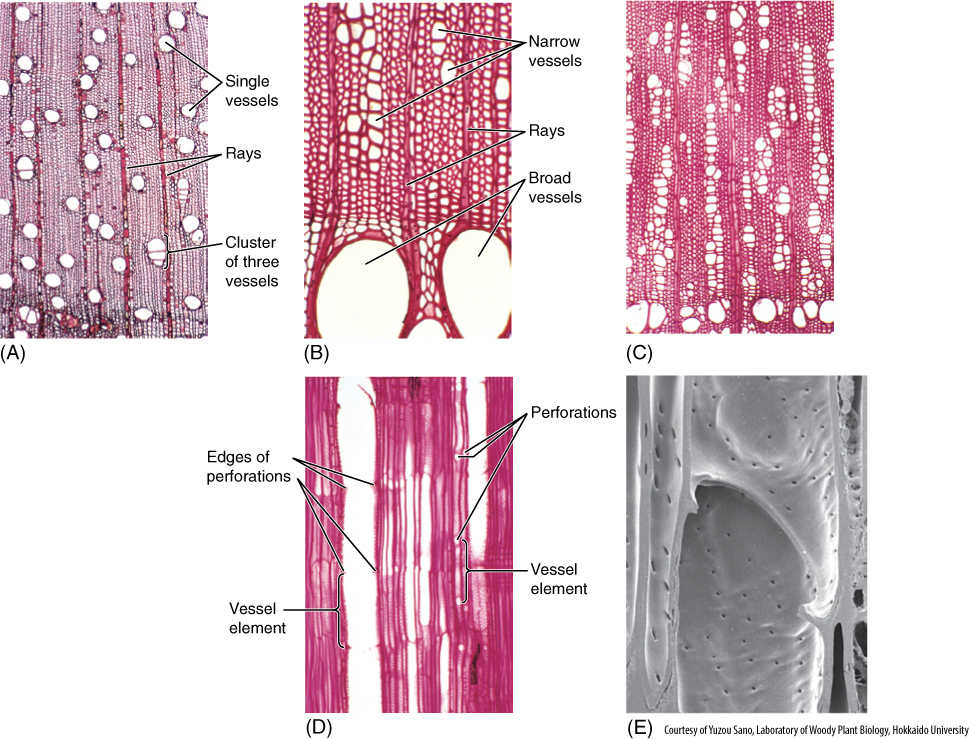
FIGURE 8-12 Vessels in wood. (A) In this wood, rather medium-size vessels occur in a mass of fibers. Each vessel occurs by itself or in a small cluster of two or three vessels (transverse section; ×40). (B) This wood has vessels of two very different sizes: extremely broad (at bottom of the micrograph) and very narrow (near the top). When water is plentiful, it can be transported in large quantities through the broad vessels. The narrow vessels are more effective at carrying water when the soil is dry and only a small amount of water is available to be transported (Cotinus americana, transverse section; ×150). (C) Vessels of Pistasia mexicana are arranged in long radial groups. Because most vessels touch two or three other vessels, they can share water through pits on their side walls (transverse section; ×50). (D) This longitudinal section passes through the lumen of one of the vessels. The front and back walls of the vessel elements were cut away during specimen preparation, and thus, the vessel appears empty. The perforations are visible as short projections into the lumen (radial section; ×150). (E) Simple perforation between two vessel elements: The front portion of each has been cut away and we are looking at the back half of the two elements. Water would have moved from the lower cell to the upper cell through the large hole, the perforation. There are numerous small pits in the side walls (×3,000).
The axial system always contains tracheary elements (tracheids or vessel elements or both), which carry out longitudinal conduction of water through the wood (FIGURE 8-12). In many species of woody angiosperms, the axial system also contains fibers that give the wood strength and flexibility (FIGURE 8-13 and TABLE 8-2). Most commercially important angiosperm woods contain large amounts of fibers, making them strong, tough, and useful for construction. They are called hardwoods, a term now used for wood of all basal angiosperms and eudicots, even those that lack fibers or are very soft, such as balsa. Woods from conifers such as pines and redwoods have few or no fibers and thus have a softer consistency. These are known as the softwoods, even though in many instances (such as bald-cypress, Taxodium) they are actually much harder than many hardwoods.
Tracheary elements and fibers are elongate, as are fusiform initials that produce them, but in many species, some immature cells undergo transverse divisions and differentiate into columns of xylem parenchyma (FIGURE 8-14). This is axial xylem parenchyma, and it is important as a temporary reservoir of water; on cloudy or humid days and at night when leaves are losing little water, wood has a temporary surplus of conducting capacity, and water is moved from roots into wood parenchyma and held there. Later, when the air is hot and dry, leaves may lose water very rapidly; water can then be drawn from wood parenchyma if the soil is too dry to supply enough water. Many desert-adapted trees have abundant xylem parenchyma. In contrast, conifers such as pine, cedar, juniper, and redwood have little or no axial parenchyma and therefore have little reserve water. For them, tough, waxy, water-conserving leaves are selectively advantageous.
TABLE 8-2 Cell Types Present in Wood
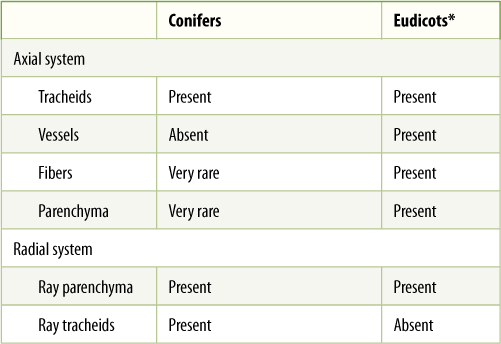
*Eudicot wood is extremely variable; certain species lack some of these cell types, other species have them all. The relative amounts of each vary greatly.
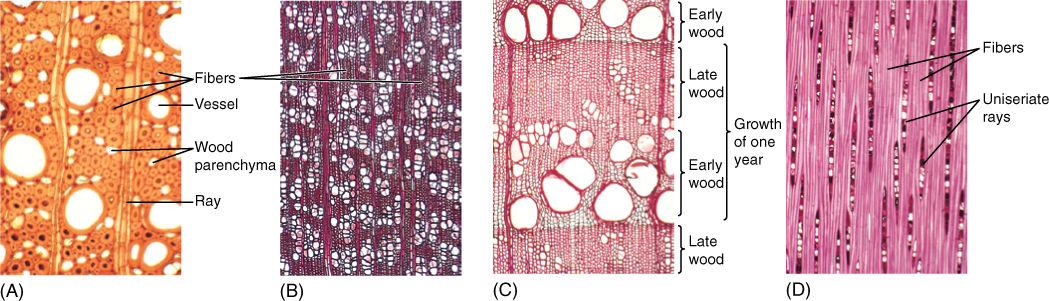
FIGURE 8-13 Wood fibers. (A) The fibers of this wood have extremely thick walls. Also, fibers are abundant, so this wood is quite strong (transverse section; ×150). (B) Vessels make up over half the volume of this wood, and thus, fibers are not very abundant. Furthermore, the fiber cell walls are rather thin; this would not be a strong wood (transverse section; ×40). (C) In this wood, regions that have many large vessels and few fibers (called early wood, discussed later in this chapter) alternate with regions that have abundant fibers and only a few narrow vessels (late wood). Early wood provides maximum conduction capacity, whereas late wood provides strength (transverse section; ×50). (D) A longitudinal section through fiber-rich late wood. All fiber cells are long and narrow and have thick walls (tangential section; ×50).
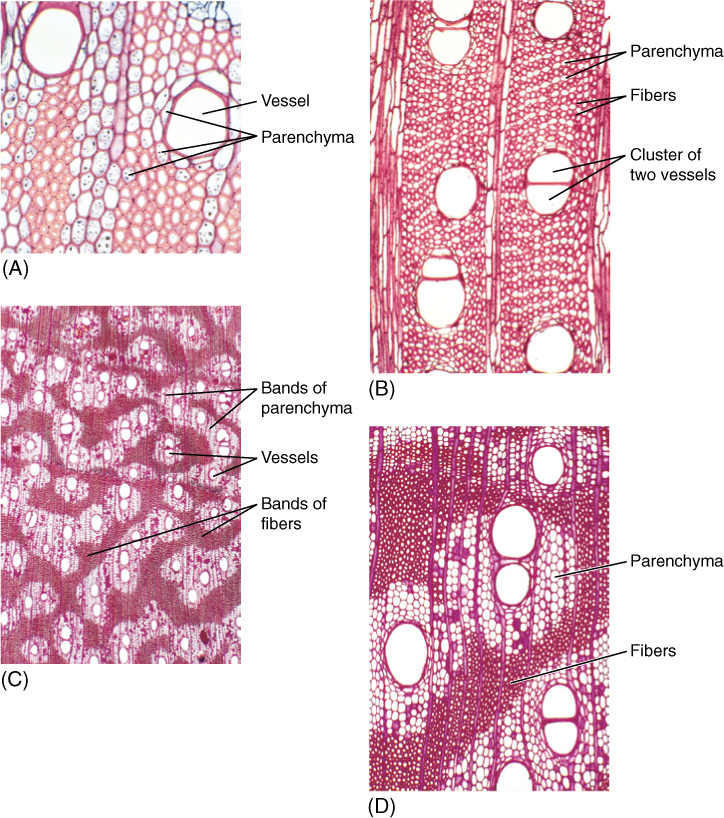
FIGURE 8-14 Wood axial parenchyma. (A) To prepare wood for microscopy, it is often boiled to make it soft enough to cut into thin specimens; unfortunately, this destroys the protoplasm of any parenchyma cells. This wood was not boiled, so the starch-filled parenchyma cells are easily visible. The large cells surrounding the vessels are also parenchyma, and they can release water to the vessels in times of water shortage (temporarily dry soil) (transverse section; ×150). (B) This wood has abundant axial parenchyma—the small cells with thin walls. The small cells with thick walls are fiber cells. Because this wood was boiled, it is difficult to be certain which is parenchyma (transverse section; ×50). (C) and (D) The axial parenchyma in this wood forms large bands; all of the vessels occur in the bands, surrounded by parenchyma. None is in the fiber masses. This parenchyma may act as a “water jacket” around vessels, absorbing excess water when water loss from leaves is low (cool nights) and releasing it when water loss is rapid (hot days) (transverse sections; (C) ×15; (D) ×50).
The complexity of the axial system of wood varies greatly. Most conifers contain only tracheids in their axial systems; fibers and parenchyma cells are sparse or absent (see Table 8-2). Some angiosperms, especially some basal angiosperms, have mostly just tracheids; however, in the majority of woody angiosperms, all possible types of cells are present, and numerous cell-cell relationships are possible. Water-storing parenchyma may be immediately adjacent to vessels, or it can be arranged such that it never touches vessels. Fibers provide maximum strength if grouped together in masses, which is how they are usually arranged (Figure 8-14D). If fibers are located around a vessel, their secondary walls reinforce the walls of the vessel and help it resist collapse, but the presence of fibers excludes parenchyma cells.
The radial system of xylem is usually simple. In woody angiosperms, it contains only parenchyma, arranged as uniseriate, biseriate, or multiseriate masses called rays (Figure 8-11 and FIGURE 8-15). Ray parenchyma cells store carbohydrates and other nutrients during dormant periods and conduct material over short distances radially within wood. The two basic types of ray parenchyma cells are upright cells and procumbent cells (Figure 8-15). At least in some plants, procumbent ray cells have no direct connection with axial cells, but upright cells do. The ray/axial interface can take many forms. If the upright ray parenchyma cell is adjacent to axial parenchyma, plasmodesmata occur. If the ray parenchyma is adjacent to an axial tracheid or vessel element, the tracheary element has pits in its secondary wall and the ray cell has very thin walls facing the pits. In early springtime, when trees such as maples are drawing on their nutrient reserves, the starch that had been stored in the upright cells is the first to be digested into sugar and passed into the axial tracheary elements for conduction to newly expanding buds, leaves, and flowers. Starch in procumbent cells is not digested until later and presumably must first be routed through upright cells for transfer to axial conducting cells.
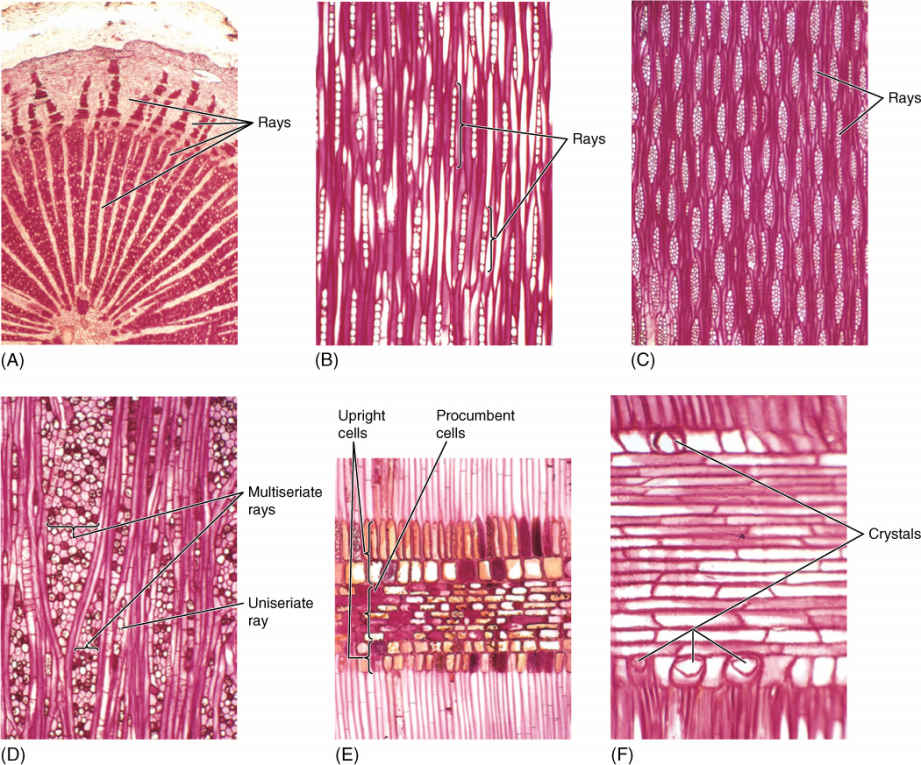
FIGURE 8-15 Wood rays. (A) The rays in this wood are extremely broad, and thus, they are easy to see. Each ray extends from the xylem into the phloem (×15). (B) These rays are uniseriate, only one cell wide; they are rather short, only about 10 cells tall. This is from late wood, so vessels are not present in this section (tangential section; ×50). (C) These rays are multiseriate, several cells wide. Over half the volume of this area of the wood is storage parenchyma, not conductive tracheary elements or strengthening fibers (tangential section; ×40). (D) This wood has giant multiseriate rays and small uniseriate rays (tangential section; ×50). (E) This ray has procumbent cells in the central part and upright cells along the edges (radial section; ×50). (F) Ray cells often contain crystals of stored material; four crystals are visible here in the cells along the margin. None occurs in the procumbent cells (radial section; ×150).
In conifers, xylem rays are almost exclusively uniseriate; they are multiseriate only if they contain a resin canal (FIGURE 8-16A). In addition to ray parenchyma cells, they may contain ray tracheids—horizontal, rectangular cells that look somewhat like parenchyma cells but have secondary walls, circular bordered pits, and protoplasts that degenerate quickly after the secondary wall is completed (FIGURE 8-16B).
Growth Rings
In regions with strongly seasonal climates, the vascular cambium is quiescent during times of stress, either winter cold or summer drought, but when quiescence ceases, the vascular cambium becomes active and cell division begins. At the same time, the new, expanding leaves are thin and delicate, and their cuticle is neither thick nor fully polymerized. Leaves like this lose water at a rapid rate, and thus, trees need a high capacity for conduction at this time. The first wood formed is early wood, also called spring wood, and it must have a high proportion of wide vessels (FIGURE 8-17A and 8-17B) or, in conifers, wide tracheids (FIGURE 8-17D). Later, the cuticle has thickened, transpiration is less, and large numbers of newly formed vessels are conducting rapidly. Wood produced at this time, called late wood or summer wood, can have a lower proportion of vessels. But the plant is a year older, is larger and heavier, and it needs more mechanical strength to hold up the increased number of leaves and the larger branches. Late wood is stronger if it contains numerous fibers or, in conifers, if it contains narrow, thick-walled tracheids. Finally, at the end of the growing season, the cambium becomes dormant again. The last cells often develop only as heavy fibers with especially thick secondary walls. In a tree with wood like that just described, it is easy to see early wood and late wood, the two together making up 1 year’s growth, an annual ring. If a summer is unusually cold, a tree may fail to grow and produce a ring, so these are occasionally not truly annual rings. Some people prefer the term growth ring.
An alternative arrangement exists: In some species, vessels form throughout the growing season. Those produced in spring are neither more abundant nor obviously wider than those produced in summer. Because the wood of a growth ring has vessels located throughout it, it is said to be diffuse porous (FIGURE 8-17C), whereas species with vessels restricted mainly to early wood are ring porous. Examples of trees with diffuse porous wood are yellow birch, aspen, sugar maple, and American holly; trees with ring porous wood include red oak, sassafras, and honey locust. In mild tropical climates, the cambium may remain active almost continuously, and the wood of one year is difficult to distinguish from that of another; growth rings are indistinct.
Heartwood and Sapwood
The center of a log is almost always darker in color than the outer wood, and it is usually drier and more fragrant (FIGURE 8-18). The dark wood is heartwood and the lighter, moister outer region is sapwood. The different regions exist because vessels and tracheids do not function forever in water conduction; water columns break because of freezing, wind vibration, tension, wood-boring insects, and other factors. After the water column breaks, there is no means of pulling water upward; vessels and tracheids in which this has occurred usually never conduct water again. Although only a few water columns break at any time, after several years, all water columns in a growth ring have snapped, and that ring no longer conducts. New water-filled tracheary elements are produced by the cambium during the next year. An important problem is that a vessel is wide enough that a fungus can easily grow up through it; for vessel elements that are not conducting, a mechanism that seals them off is selectively advantageous. Wood parenchyma cells adjacent to vessels push bubbles of protoplasm through the pits into the vessel, forming a plug, called a tylosis (plural: tyloses), completely across it (FIGURE 8-19). This occurs repeatedly, and the vessel may become filled. These and other wood parenchyma cells undergo numerous metabolic changes and produce large quantities of phenolic compounds, lignin, and other dark-colored, aromatic substances that inhibit growth of bacteria and fungi. These chemicals are usually dark and aromatic, and as they accumulate, wood becomes darker and more fragrant, such as cedar wood. Ultimately, all parenchyma cells die, and conversion of sapwood to decay-resistant heartwood is complete.
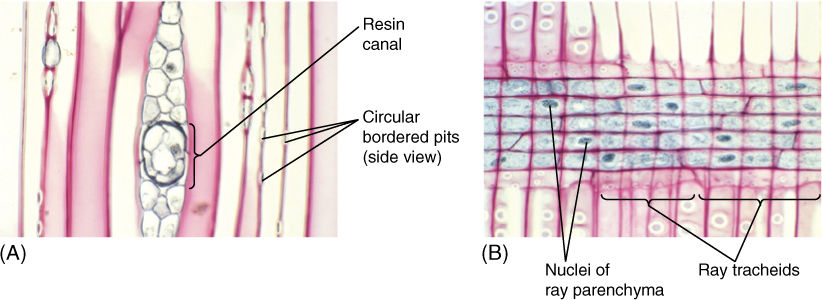
FIGURE 8-16 (A) Conifer rays often contain secretory canals that produce resin (pitch), important in preventing insects from burrowing through the wood (tangential section; ×150). (B) The central ray cells are living ray parenchyma, with large nuclei that have been stained black. The procumbent cells along the top and bottom of the ray are ray tracheids that have circular bordered pits but no protoplasm when mature (radial section; ×150).
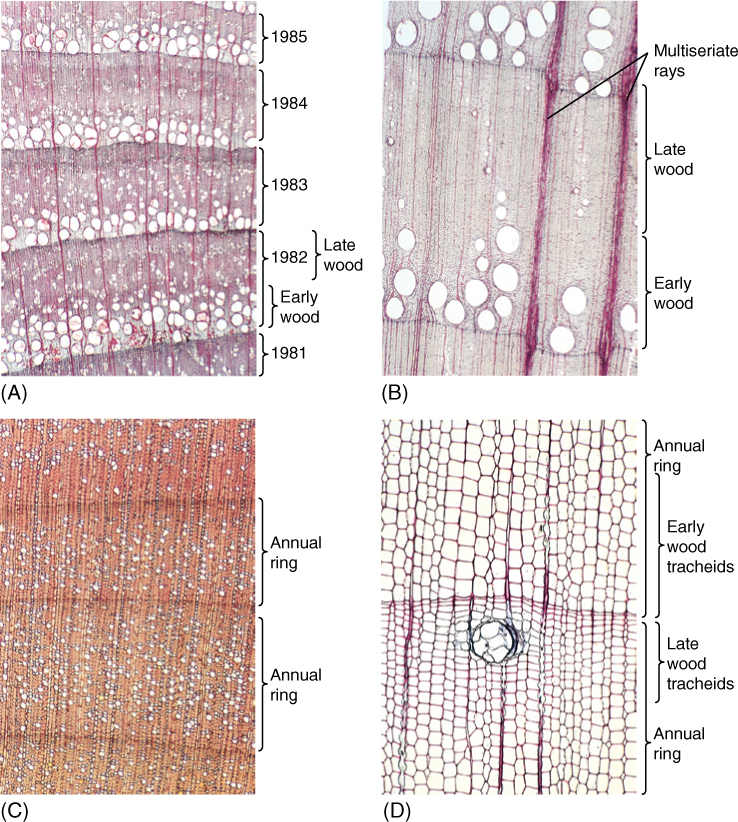
FIGURE 8-17 (A) In this transverse section, portions of five growth rings are visible. The lowermost ring is oldest; the next higher ring was formed 1 year after the lowest ring, and so forth. The vascular cambium is located beyond the top of the photograph. The wood is ring porous, and vessel-rich early wood is easily distinguished from fiber-rich late wood (transverse section; ×50). (B) This wood is very strongly ring porous, with large vessels formed only when the cambium first becomes active. Later, the cambium produces fibers exclusively, except for a few rare, very narrow vessels. Two large multiseriate rays are visible, but most rays are narrow and uniseriate (transverse section; ×15). (C) This wood is diffuse porous, with vessels occurring rather uniformly in both late wood as well as early wood. Growth rings are not as conspicuous as in ring porous wood (transverse section; ×40). (D) Most conifer wood has only tracheids in its axial system, but it still has early wood (wide tracheids) and late wood (narrow tracheids) (transverse section; ×50).
A new layer of sapwood is formed each year by the vascular cambium, and on average, one annual ring is converted to heartwood each year. Thus, whereas heartwood becomes wider with age, sapwood has a more or less constant thickness (TABLE 8-3). Of course, this is not true of young stems and roots: Black walnut typically has wood that functions for 10 years before converting to heartwood. In a seedling or a branch only 9 years old, no heartwood is present yet.
Plants Do Things Differently
BOX 8-2 Having Multiple Bodies in One Lifetime
Woody plants have two bodies. As the primary body of a woody plant ages, a vascular cambium arises inside it and produces wood and secondary phloem—an entire new body—inside the preexisting body. Think about how different the two bodies are. The primary body has leaves and axillary buds, flowers, fruits, and seeds. It has root hairs and absorbs water and nutrients. The secondary body is just wood and bark. It has no leaves, no buds, no flowers, and so on. The secondary body is, for the most part, nothing more than an ever-growing vascular/skeletal system. The two bodies look completely different and have distinct functions. Growth of the secondary body tears apart and destroys the primary phloem, cortex, and epidermis of the plant’s primary body, and these dead remnants are shed as part of the plant’s first bark. Shoot tips and root tips continually make more sections of primary body, but they too will be destroyed by formation of more secondary body inside them. It is dramatic for one organism to have two distinct bodies, to have one body form inside another, destroying the first. Does anything like this occur in animals? In us? Yes, and in even more dramatic fashion.
We humans undergo moderate changes in our body. When about 5 or 6 years old, we shed our baby teeth as a new set of permanent teeth forms below them. As the permanent teeth develop and enlarge, they simply push our baby teeth out and we lose them. Parts of our body—teeth with blood vessels, nerves, and living cells—just fall out much the way bark falls off a tree. Later, when we go through puberty, other changes occur. Hair follicles, especially in boys, become active and start producing thicker hair than the type children have. In girls, there is development of glandular and adipose tissue in the breasts. These and other changes, however, are really just modifications of preexisting tissues that were already present in children. There is nothing equivalent to a cambium and the formation of brand new cells.
Our puberty, however, pales in comparison to that of eels. Juvenile eels are just tiny, flat, coin-shaped marine fish that look something like a leaf. As they go through the transformation to being adults, they develop their very long, cylindrical shape and switch to being freshwater fish, migrating up rivers to spawn. Juvenile and adult eels have such different bodies that the juveniles were long considered to be a completely different type of fish. Juvenile and adult Homo sapiens are obviously the same species. We do not change that much during puberty.
Other animals go through more significant bodily changes. Crabs, lobsters, and beetles have an exterior exoskeleton that is so hard it cannot grow. The animal periodically produces a new, soft exoskeleton; then the animal molts—that is, it sheds its skin and old exoskeleton—and very quickly grows to a new size before its new exoskeleton hardens and prevents further growth. After some time, the animal will repeat this process so that it can grow even larger. Snakes too periodically shed an old skin, replacing it with a new one. In these examples, entire, complex tissues are being sloughed off and new body parts are formed.
Undoubtedly, the most drastic examples of individuals that have two distinct bodies are insects that go through what is called a complete metamorphosis. Their larval bodies do not look anything at all like the adult bodies. Examples are caterpillars, which metamorphose into moths or butterflies, and maggots, which metamorphose into flies. The larval body is specialized for eating and growing and has neither wings nor sex organs of any kind. In contrast, the adult body does have wings and sex organs, enabling it to fly about and find a mate and then carry out sexual reproduction and dispersal. In some cases, the adult body needs to survive only a day or two until it can mate and die (males) or lay fertilized eggs (females). Such adult bodies have either no digestive system at all or such a simple one that it can only absorb the sugar water of flower nectar. In the complete metamorphosis of caterpillars, the caterpillar spins a cocoon around itself; then its body more or less dissolves except for special sets of cells called imaginal discs. These act like meristems and produce the cells, tissues, and organs of the adult body by using the nutrients from the dissolved larval body. By the time metamorphosis is complete, the body has been completely rearranged; almost nothing exists of the preexisting larval body except that its molecules have been recycled and not wasted.
A plant’s primary body differs as greatly from its secondary body as do the caterpillar body and the butterfly body of a particular species. If plants could also digest their primary bodies and rebuild them the way caterpillars do, the transformation in plants would be seen to be just as dramatic as metamorphosis in animals. However, plants do not form cocoons and do not undergo self-digestion, so the activity of a vascular cambium and a cork cambium in the production of an entire new secondary body seems unremarkable. It appears as if the plants are doing nothing more than adding a few new tissues, but the change is really much more fundamental.
Just considering ourselves, the idea of individual plants and animals having several distinct bodies may seem far fetched, but as it turns out, it is a common occurrence with each body carrying out distinct phases of the organism’s life activities.

FIGURE 8-18 (A) Transverse section of a water oak trunk (Quercus nigra). The outermost thin brown band is the bark. The broad bands of yellowish tissue and the innermost band of brown tissue are wood. The yellowish wood (sapwood) is young, and many of its cells are still alive and many of its vessels were still conducting water when the tree was cut down. The brownish wood (heartwood) is the oldest, and all its cells had died and all its vessels had cavitated. Most of the cells in wood have thick secondary walls that are heavily lignified; consequently, wood cells decompose slowly. Many trees live for hundreds of years, so the carbon dioxide they capture (sequester) and use to build wood is kept out of the atmosphere for hundreds of years. Trees also absorb unusual chemicals in the air and water and lock them into the tree rings. By carefully cutting out just a bit of a tree ring, we can analyze many aspects of the environment as it existed in the past. (B) High magnification view of the vascular cambium, sapwood, and outer heartwood shown in (A). Vessels are wide enough to be visible. Both images are water oak, Quercus nigra.
TABLE 8-3 Thickness of Sapwood in Eudicot Trees
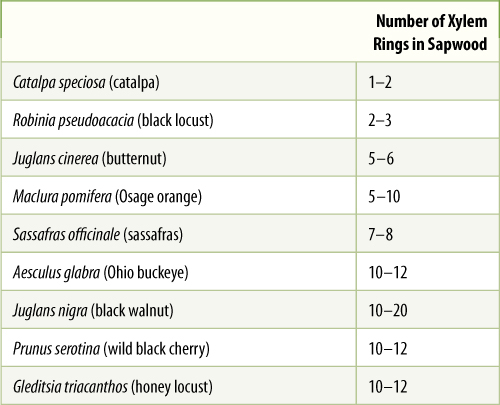
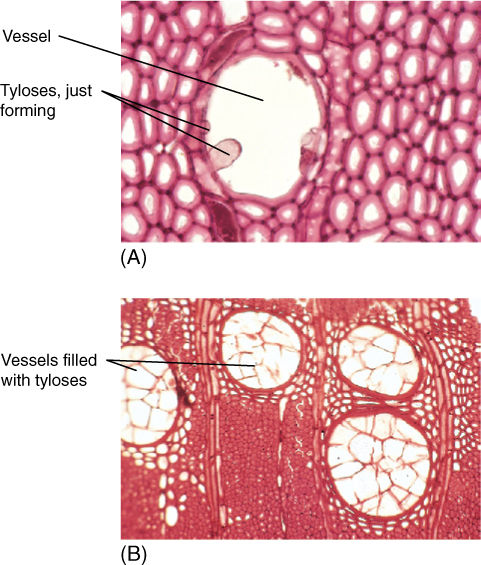
FIGURE 8-19 (A) Tyloses form as protoplasm from surrounding parenchyma cells pushes into a vessel. This was just beginning to form tyloses when the wood was collected for microscopy. Boiling destroyed the protoplasm (transverse section; ×250). (B) The vessels of this wood are completely occluded by tyloses; this sample came from a piece of heartwood (×50).
Reaction Wood
In branches or trunks that are not vertical, gravity causes a lateral stress; if not counteracted, the branch would droop and become pendant. In response to such stress, most plants produce reaction wood. In angiosperms, this develops mostly on the upper side of the branch and is known as tension wood. In a cross-section of such a branch, growth rings are eccentric, being much wider on the top of the branch. Tension wood contains many special gelatinous fibers whose walls are rich in cellulose but have little or no lignin. These fibers exert tension on the branch, preventing it from drooping, or the tension wood may even contract, slowly lifting a branch to a more vertical orientation. Conifers form reaction wood located on the underside of the branch and is known as compression wood. It is enriched in lignin and has less cellulose; growth rings are especially wide on the lower side of the limb.
![]() Secondary Phloem
Secondary Phloem
Because secondary phloem is formed from the vascular cambium just as secondary xylem is, it too has an axial and a radial system (FIGURE 8-20). The axial system is responsible for conduction up and down the stem or root; it contains sieve tube members and companion cells in angiosperms, or sieve cells in conifers. In both groups of plants, fibers and nonconducting parenchyma are also typically present in axial secondary phloem. In some species, there may be bands of fibers alternating with sieve tube members, but usually, these are not annual rings, and many other patterns can be seen in other species. Whereas the equivalent cells of axial secondary xylem are arranged as early and late wood and as ring porous or diffuse porous wood, no similar arrangement occurs in secondary phloem. Although the tracheary elements of secondary xylem may function for many years before being converted to heartwood, sieve tube members and sieve cells usually conduct for less than 1 year; only the innermost layer of phloem is capable of conduction.
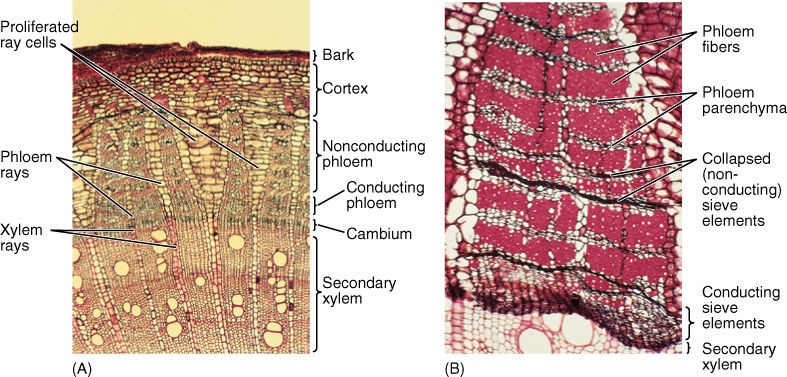
FIGURE 8-20 (A) This stem transverse section has secondary xylem at the bottom, a vascular cambium region, secondary phloem, and then cortex and bark at the top. Xylem rays and phloem rays meet at the cambium. Parenchyma cells of some rays have proliferated through cell division and expansion, which has prevented tearing of the bark (×40). (B) In this secondary phloem of Artabotrys, only the youngest axial phloem (at the bottom) contains functional sieve tube members. The youngest phloem and the vascular cambium are so soft that they are often damaged while trying to cut samples from trees; this is why they are partially crushed here. Older phloem has abundant fibers alternating with bands of phloem parenchyma and collapsed sieve tube members (×50).
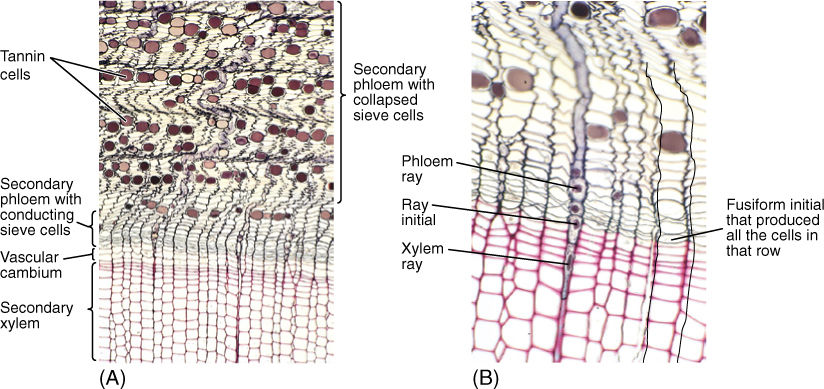
FIGURE 8-21 (A) and (B) The outermost, youngest wood and the bark of pine. As sieve cells stop functioning and collapse, phloem shrinks and becomes undulate. This causes rays to become wavy. Phloem rays meet xylem rays at the ray initials. Also, each row of sieve cells was produced by the same fusiform initial that produced the corresponding row of tracheids (transverse sections; (A) ×50; (B) ×150).
The size, shape, and number of phloem rays match those of xylem rays because both are produced by the same ray initials (FIGURE 8-21). Phloem rays consist only of parenchyma cells that are used for storage, as are xylem rays, but phloem rays seem to be even more important for this (FIGURE 8-22). In Conifers, albuminous cells are ray cells.
![]() Outer Bark
Outer Bark
Cork and the Cork Cambium
The production and differentiation of secondary xylem cells cause the vascular cambium and secondary phloem to be pushed outward. As the youngest, innermost phloem cells form and mature, they contribute to the larger diameter of the stem or root and increase pressures acting on the outermost tissues. This requires that tissues on the periphery of the plant either grow in circumference or be torn apart. Actually, the tissues do both, but both must be controlled (Figure 8-20). The integrity of the plant surfaces must be maintained against invasion by fungi, bacteria, and insects. As circumferential stretching increases and the older sieve elements die, some storage parenchyma cells become reactivated and undergo cell division. This is similar to the activation of parenchyma cells during formation of the interfascicular vascular cambium, but in secondary phloem, it results in a new cambium, the cork cambium, also called the phellogen (FIGURE 8-23).
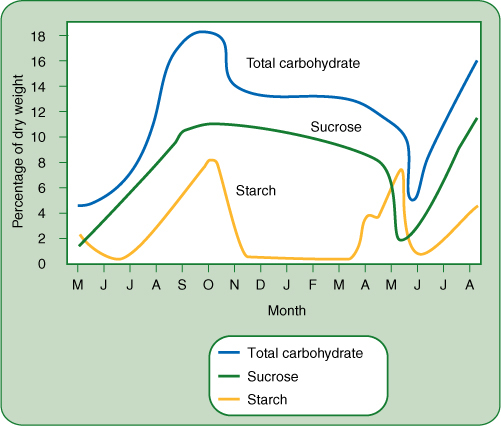
FIGURE 8-22 Carbohydrate accumulates in the bark of stems and roots during early summer while leaves are still present and photosynthesizing; it reaches its peak in September and October, then drops and remains steady through winter (December to April). It is released during the spring growth season, April and May. These data are for black locusts near Ottawa, Canada, where spring comes late. For plants that grow farther south, the spring release occurs earlier.
Cork cambium differs greatly from vascular cambium in both structure and morphogenic activity. All of its cells are cuboidal, like ray initials. After each division, the inner cell almost always remains cork cambium, whereas the outer cell differentiates into a cork cell (also called a phellem cell). In a few species, the cork cambium may produce a cell or two to the inside that mature into a layer of parenchyma called phelloderm. The cork cambium, the layers of cork cells, and the phelloderm (if any) are known as periderm. Maturing cork cells increase slightly in volume; then their thin primary walls become encrusted with suberin, making them waterproof and chemically inert, and then they die. Cell death is probably a critical part of maturation because the protoplasm breaks down, leaving nothing digestible or nutritious for an animal to eat. In many species, some cells deposit secondary walls and mature into lignified sclereids; these usually occur in layers that alternate with cork, resulting in a periderm that is both impervious and tough (FIGURE 8-24). Because periderm is such an impermeable barrier, all plant material exterior to it, such as epidermis, cortex, and older secondary phloem, dies for lack of water and nutrients.
Periderm offers only temporary protection because the root or stem continues to grow interior to it, pushing it outward and stretching it circumferentially. Unlike vascular cambium, cork cambium is typically short lived; it produces cells for only a few weeks, after which all cells differentiate into cork cells and die. The layer of cork cells cannot expand much circumferentially, and after one or several years, a new cork cambium must be formed in younger secondary phloem closer to the vascular cambium. These new cork cells act as a further barrier and also block water and nutrients from reaching any secondary phloem cells located between layers of cork cambium. In this fashion, several layers of cork can build up.
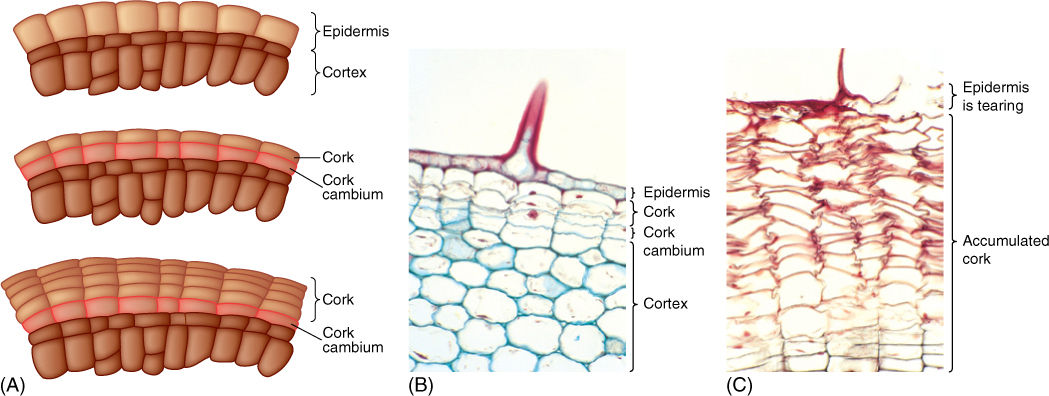
FIGURE 8-23 (A) The first cork cambium may form as epidermis cells resume mitotic activity. All divisions are by periclinal walls, and the inner cell continues as cork cambium while the outer cell differentiates into cork. (B) Hypodermis cells have just started to undergo cell division, resulting in formation of a cork cambium. This is a young stem of geranium; notice the base of a trichome (×150). (C) Older stem of geranium; many layers of phellem (cork cells) have accumulated owing to cork cambium activity. The phellem cells are dead and empty. A trichome was present here also, and formation of bark blocked transfer of nutrients to the trichome and other epidermal cells, killing them (×150).
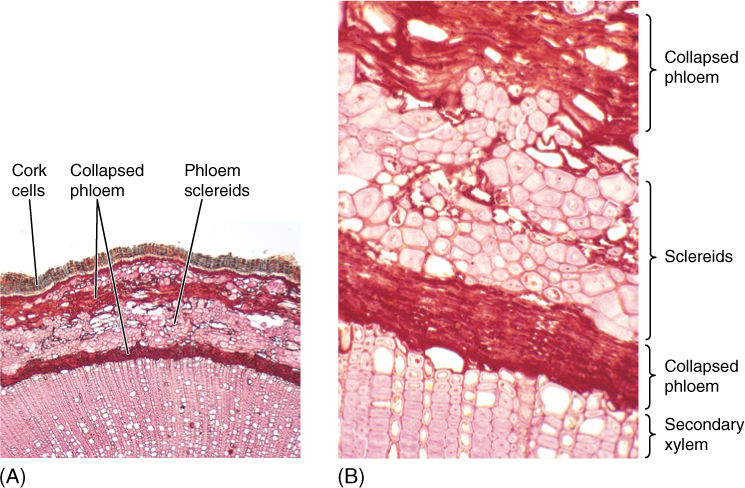
FIGURE 8-24 (A) A thick layer of uncollapsed cork cells is present on this stem. Many parenchyma cells in the cortex and secondary phloem have converted to sclereids (B), making the bark stronger and more protective. The next cork cambium will form deep in the secondary phloem, where numerous parenchyma cells are capable of becoming mitotically active again; sclereids cannot resume cell division (transverse sections; (A) ×40; (B) ×150).
All tissues outside the innermost cork cambium comprise the outer bark (FIGURE 8-25). All secondary phloem between the vascular cambium and the innermost cork cambium is the inner bark. The amount of bark is quite variable from species to species; in some, only a small amount of cork is formed, so the bark is thin and consists mostly of dead secondary phloem. In others, cork is produced in large amounts and becomes 3 or 4 cm thick. Although it is usually not obvious, bark is continuously falling off the tree, but it does not accumulate at the base because wind and water carry it away.
Lenticels and Oxygen Diffusion
The impermeability of cork has negative as well as advantageous consequences. Although it keeps out pathogens and retains water, it also blocks absorption of oxygen, interfering with the respiration of the sapwood, vascular cambium, and inner bark. Bark becomes permeable to oxygen when cork cambium produces cork cells that become rounded as they mature. Because rounded cells cannot fit tightly together, intercellular spaces penetrate the cork layer, creating a diffusion pathway for oxygen. These regions of aerenchymatous cork are lenticels (FIGURE 8-26). When a new cork cambium arises interior to this one, it too forms a lenticel in the same place; the outer and inner lenticels are aligned, permitting oxygen to penetrate across all layers of the bark. Lenticel-producing regions of cork cambia are more active than adjacent regions that produce only ordinary impermeable cork; consequently, lenticels contain more layers of cells and protrude outward. In species that have smooth bark, even small lenticels are easy to identify. On plants that have thick rough bark, lenticels can be almost impossible to see, but generally, they are located at the bases of the cracks in the bark. When the bark of cork oak is made into bottle corks, it is necessary to cut them so that the lenticels do not run from the top of the cork to the bottom.

FIGURE 8-25 (A) As this pine trunk increased in diameter, its bark was stretched and ultimately cracked. The deepest bark (still brown) is youngest, that on the surface is oldest. A forest fire charred and blackened the outermost layers of bark, which were already dead, as are the outermost layers of bark of all trees. The bark acted as insulation and prevented the fire’s heat from damaging living tissues interior to the bark. (B) Bark of maple peels off in large thin sheets because numerous cork cambia form close together and each is sheetlike. (C) Cork cells of sycamore contain many chemicals; as outer patches of bark peel away, fresh patches are exposed. When their pigments oxidize, they turn gray. Each cork cambium forms as a small patch; the size and shape of the cork cambia affect the nature of the bark. (D) The bark of cork oak (Quercus suber) becomes extremely thick and is composed mostly of phellem, with few sclereids. Cork oaks grown in Spain and Portugal provide most commercial corks used to seal bottles. When the bark has become sufficiently thick, the outer bark is peeled away; after a few years, the bark is once again thick enough to harvest. (E) This is cinnamon bark; we grind it into a powder to use as a spice. It is one of the few barks we eat.
Plants and People
BOX 8-3 Dendrochronology—Tree Ring Analysis
The amount of wood produced by the vascular cambium is closely correlated with climate. Species that occur in extremely harsh climates produce very little wood, often only a single layer of vessels each year. Slow growth such as this occurs in cold regions at high altitudes or high latitudes or in hot, dry desert zones. In the tropics, however, the amount of growth each year can be large, with each growth ring being several millimeters wide and consisting of 50 to 100 layers of new cells. Even within a single plant, vascular cambium produces more wood in an optimal year than it does in a year with poor temperatures or too little rain. This can be seen easily in the varying width of tree rings (FIGURE B8-3).
Because growth rings reflect climate so accurately, they are used for many critical studies. Starting from the most recent, outermost ring, it is possible to count toward the center of the tree, encountering older and older rings. Not only can the age of the tree be determined, but climatic fluctuations can be inferred from the width of the rings. The oldest living trees of a region can be studied for climatic changes that occurred during the lifetime of the tree, which in some individuals is more than 5,000 years. However, the analysis of past climates can be extended beyond the age of the oldest living trees. Historical or archaeological sites may contain houses, bridges, or ships constructed with large wooden beams. Some of the outermost rings of those beams may match the patterns of the inner rings of living trees. By counting back and finding the age of that set of rings in the living wood, we can determine how old that portion of the beam is and therefore when the house, ship, or bridge was constructed. This technique has been valuable not only in dating ancient settlements but also in establishing whether people lived in times of good or poor climate.

FIGURE B8-3 Some growth rings in this wood are much wider than others. The innermost ring marked “Good year” is very wide, with abundant early wood (the lighter, inner part) as well as late wood (the darker, outer part). Conditions must have been very good for the tree that year: It might have been a summer of optimal temperatures or rain; if a cultivated tree, it might have been fertilized that year; or surrounding trees that had been shading it either might have died or been cut away. The next two growth rings, both marked “Poor year,” were produced when the tree was not vigorous enough to make very much wood.
After a tree ring sequence has been mapped out from many different sources of wood, it is important to establish an exact date to the rings. In North America and much of Europe, there are extensive forests with trees old enough to allow us to analyze tree ring sequences back for a thousand years or more. Rather recent archaeological sites yield wood samples that can be matched to the centers of living trees, extending the accurately dated sequences back even farther; then samples from older archaeological sites can be added to extend the sequence back in time, knowing the exact year each ring was produced. Even if we find a wooden post used in some ancient building or ship, we can match the ring sequence to the known sequence and determine exactly when the tree lived and when it was probably cut down to be used.
In the Middle East, in areas where many critically important civilizations flourished, such as Egypt, Babylon, and Sumer, it has only been possible to map out the tree ring sequence from the present back to CE 362 (CE is “Common Era,” an alternative to AD); then we run into a lack of the necessary archaeological wood samples. By going back only to the fourth century CE, we can determine exactly when many Byzantine buildings were constructed but nothing earlier. There is a second, excellent tree ring sequence for the Middle East, but it is a “floating” tree ring chronology. There are enough archaeological wood samples that the rings of some overlap those of others, and those overlap even others so that a well-documented sequence has been established; however, none is as recent as CE 362. None comes up to the sequence where we know exactly which ring corresponds to which year. Instead, all we know is that the floating sequence extends over 1,503 years and that it must have begun somewhere earlier than 2200 years BCE (Before the Common Era) because we know the approximate age of some of the temples and palaces that supplied the ancient wood.
It may be that we can establish the absolute time of the floating sequence. In the North American and European sequences, a very strange ring was produced in the year 1628 BCE. It is believed that a volcanic eruption produced so much dust that sunlight was blocked around the world, and the summer was so cold that frost damage occurred in many trees. Dendrochronologists have discovered that the floating sequence for the Middle East also has an unusual ring that seems to match this one. Rather than frost damage, the ring in the floating sequence is one of extra growth. It may be that the Middle East at the time was so hot and dry that the cooling caused by the volcanic dust cloud was only enough to produce extra rain and optimal growing conditions rather than frost.
If his hypothesis—that the strange ring in the floating sequence occurred in the same year as the strange ring in other, anchored sequences—is correct we can then put exact dates to the buildings, statues, and ships that the various wood samples came from.
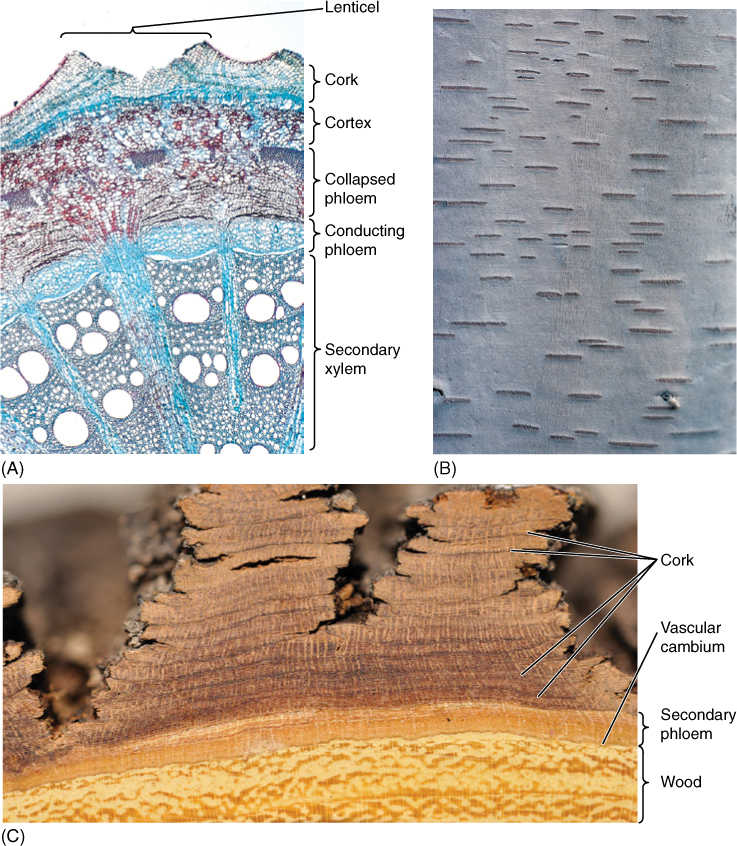
FIGURE 8-26 (A) Low magnification of stem of Aristolochia, with a large lenticel. Rapid production of cork cells in lenticels cause the epidermis to rupture. Although they cannot be seen here, small intercellular spaces allow oxygen to diffuse through the lenticel into the trunk or root (×15). (B) Lenticels of birch (Betula) widen as the trunk increases in circumference. Occasionally, they divide into two lenticels when the central cork cambium cells begin producing compact cork instead of aerenchymatous cork. (C) This bark of honey mesquite (Prosopis glandulosa) contains many small patches of cork, each produced by a cork cambium that arose from parenchyma cells in the secondary phloem, produced a few layers of cork, and then stopped functioning. Each patch of cork is surrounded by secondary phloem, and the outer patches of cork are older than the inner ones. Lenticels are not visible but would be located at the deepest part of each fissure.
Initiation of Cork Cambia
The timing of initiation of the first cork cambium is far more variable than that of the vascular cambium. In some species, the first cork cambium arises before a twig or root is even 1 year old. On stems this is often detectable as the surface color changes from green to tan. In other species, the first cork cambium forms only when that region is several years old; until then, the epidermis and cortex are retained. Epidermises more than 40 years old have been reported. Delayed formation of bark is common in plants that depend on cortex chlorenchyma for much of their photosynthesis, as cacti do. The first cork cambium may arise in a number of tissues: epidermis, cortex, primary phloem, or secondary phloem. Subsequent cork cambia may form shortly afterward, sometimes in the same season, but usually a year or two later. If the growth in diameter is slow, new cork cambia may arise at intervals of as much as 10 years. These later cork cambia usually form deep in the secondary phloem.
The first bark on young stems usually differs from bark formed when the stem is older. If the first cork cambium arises by reactivation of epidermal cells, the first outer bark contains only periderm and cuticle and is very smooth. If the first cork cambium arises in the cortex, the first outer bark contains periderm, cortex, and epidermis; this too is smooth and contains any cortical secretory cells that were present. As the first bark is shed and later cork cambia arise in the secondary phloem, they produce an outer bark that contains only cork and phloem. The nature of these later barks depends greatly on the cell types present in the trapped secondary phloem: Fiber cells produce fibrous, stringy bark, sclereid-filled phloem produces hard bark, and so on. It is not unusual for the bark of a young tree to be dramatically different from the bark it will have when it is older (FIGURE 8-27).

FIGURE 8-27 This trunk of Casuarina (red beefwood) still has fragments of its first bark, but the underlying newer bark is also visible. Young branches have only the first type of bark, older parts of the trunk only the second type.
![]() Secondary Growth in Roots
Secondary Growth in Roots
Roots of conifers and woody angiosperms undergo secondary growth, as do the stems. A vascular cambium arises just like the interfascicular cambium, when parenchyma cells located between the primary xylem and primary phloem become active mitotically, as do pericycle cells near the protoxylem (FIGURE 8-28). The new vascular cambium has the same star shape as the primary xylem, but it soon becomes round as the cambium in the sinuses of the primary xylem produces more secondary xylem than do the regions of cambium near the arms of protoxylem. Consequently, some portions of the cambium are pushed outward more rapidly than others. When a circular cambium is achieved, the unequal growth stops, and all parts grow at similar rates.
Root vascular cambium contains both ray and fusiform initials, and in many cases, wood produced in the root is quite similar to that of the shoot, having sapwood and heartwood and being ring porous or diffuse porous as the stem is. Typically, however, the wood of roots is not identical to that of stems of the same plant, and they may be totally dissimilar when the conductivity requirements of root and stem differ. For example, many cactus roots are extremely long, nonsucculent, cable-like structures. After a brief rain, the numerous root tips absorb water from a large region of surface soil. Because the soil can dry within hours after such a rain, water must be conducted rapidly and in large quantity into the succulent cactus shoot where it can be stored. Once in the shoot, however, conduction requirements are totally different; the shoot may be quite small and consist mostly of parenchyma, with very little wood and just a few narrow vessels. The water is stored so effectively that most conduction is probably by diffusion through the cytoplasm rather than by xylem. Furthermore, the narrow roots are strengthened by numerous wood fibers, whereas the small succulent shoots are strengthened only by turgor. In the shoot, the xylem and phloem rays are gigantic, providing a large region for water storage. In the roots, which do not store water, rays are small and narrow and consist mostly of sclerenchyma.
Perennial roots also form bark; the first cork cambium usually arises in the pericycle, causing the endodermis, cortex, and epidermis to be shed. The cork cambium produces cork cells to the outside, forming a protective layer, and in some species a layer or two of phelloderm as well. Lenticels also occur, being especially prominent near lateral roots. The bark on roots may be similar to that on the stem of the same plant, but in some species, there are significant differences, again related to the differing metabolisms and microhabitats of the two organs.
Several mechanisms exist by which the storage capacity of a woody root can be increased. The ray parenchyma of the secondary xylem offers considerable volume, and storage capacity is increased if the rays become larger. Axial parenchyma in the wood can also be used, and in many storage roots, wood is almost pure parenchyma (FIGURE 8-29). For example, the orange part of a carrot root that we eat is wood consisting mostly of wood parenchyma and just a few vessels. When we peel the carrot, we remove a thin bark and all secondary phloem.
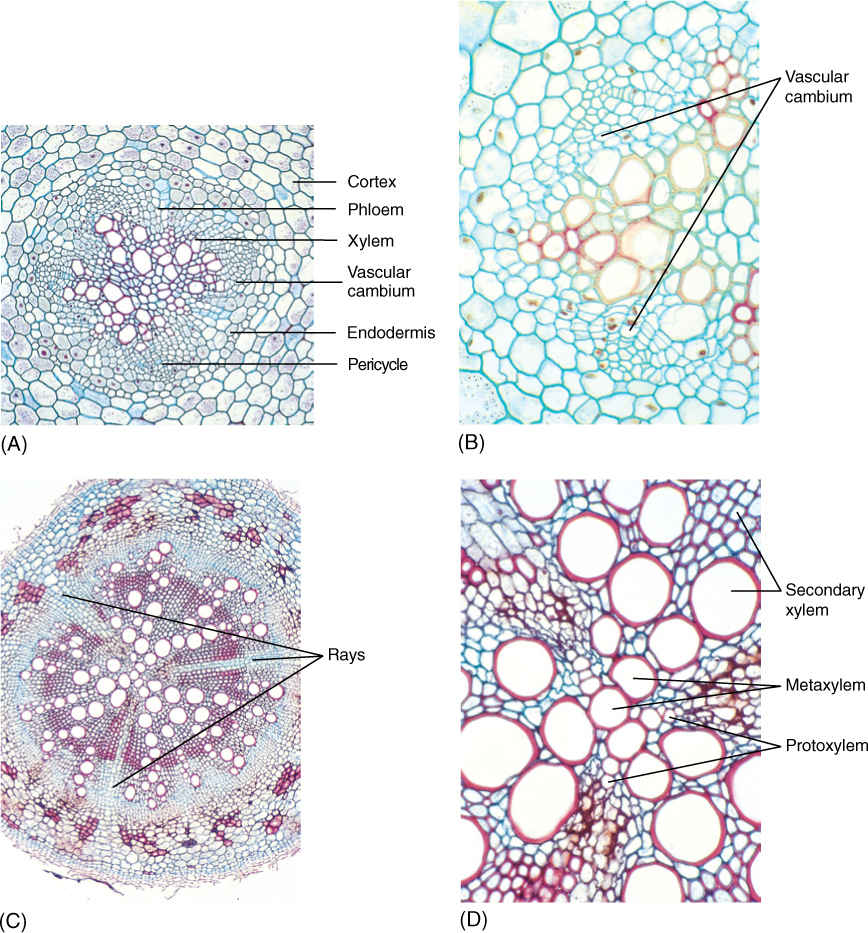
FIGURE 8-28 (A) and (B) Young roots. The vascular cambium of a root such as this baneberry arises as an undulate cylinder located around the xylem, interior to the phloem; (A) × 50; (B) × 150 (C) and (D) Older roots. The organization of root wood is similar to that of stem wood: an axial system and a set of rays. In (C), there are three large rays; the high magnification of (D) reveals that these rays are aligned with the protoxylem of the primary xylem (C) ×40; (D) ×150.
![]() Anomalous Forms of Growth
Anomalous Forms of Growth
Anomalous Secondary Growth
The development, cellular arrangement, and activity of vascular cambia in conifers and most woody angiosperms are remarkably similar. However, there are alternative types of cambium structure and activity, and analysis of the consequences of various types of arrangements reveals a great deal about secondary growth. Because alternative cambia produce secondary bodies that differ from the common type, their growth is called anomalous secondary growth.
Roots of Sweet Potatoes
In sweet potatoes (Ipomoea batatas), the amount of storage parenchyma is increased dramatically by an anomalous method of secondary growth. Numerous vascular cambia arise, not around the entire mass of primary xylem, but around individual vessels or groups of vessels (Figure 8-29B). The cambia act normally, except that the xylem and phloem produced are almost purely parenchyma. New vessels may also be surrounded by another new cambium, and the process is repeated. As the sweet potato becomes quite large, it may contain hundreds of cambia of various ages; the secondary tissues are an irregular matrix of parenchyma, a few sieve tubes, some vessels, and vascular cambia.

FIGURE 8-29 The part of a carrot (A) or radish (B) we eat is the wood of the storage root. It does not look like wood at first glance; it is mostly parenchyma with a few vessels and no fibers. This wood is well adapted for rapid production of long-term storage tissue (×40).
What is the selective advantage of producing so many vascular cambia instead of just one? It may be that the rate of cell production is important. Because the root must become large very quickly, having just one cambium may be too slow. Multiple cambia all functioning simultaneously speed the production of storage capacity.
Included Phloem
In several eudicots, a vascular cambium of the common type arises and produces ordinary secondary xylem and phloem. After a short period, however, the cambium cells stop dividing and differentiate into xylem: There is no longer a cambium. Cells in the outermost, oldest secondary phloem then become reactivated and differentiate into a new vascular cambium that acts just like the first, producing ordinary secondary xylem and phloem, then differentiating and ceasing to exist. Notice in FIGURE 8-30 that because the second cambium arose in the outermost phloem, the xylem it produces is located exterior to the phloem of the first cambium. From interior to exterior there is first xylem, first phloem, second xylem, and second phloem. Then a new vascular cambium arises in the outermost second phloem and so on. This type of secondary phloem, located between two bands of xylem, is included phloem. The selective advantage may be protection of phloem from insects and other pests by one to several layers of wood. Also, tissue relationships between xylem and included phloem differ from those between xylem and ordinary phloem, but the advantages of this have not yet been studied.
Unequal Activity of the Vascular Cambium
In ordinary growth, all areas of the vascular cambium have about equal activity, and the stem or root is therefore round in cross-section. However, in some species of Bauhinia and certain other woody vines, two sectors of the cambium are very active, while two are almost completely inactive (FIGURE 8-31). The stem grows outward in two directions but remains thin in the other two, and it soon becomes a thin, flat, woody ribbon. The selective advantage of this may be related to flexibility. As an ordinary round stem becomes wider in all directions, its conducting capacity increases, but its flexibility decreases. As the Bauhinia stem becomes wider, its conducting capacity increases, but its flexibility remains about the same. For many vines, flexibility is selectively advantageous, and this type of anomalous secondary growth is adaptive.
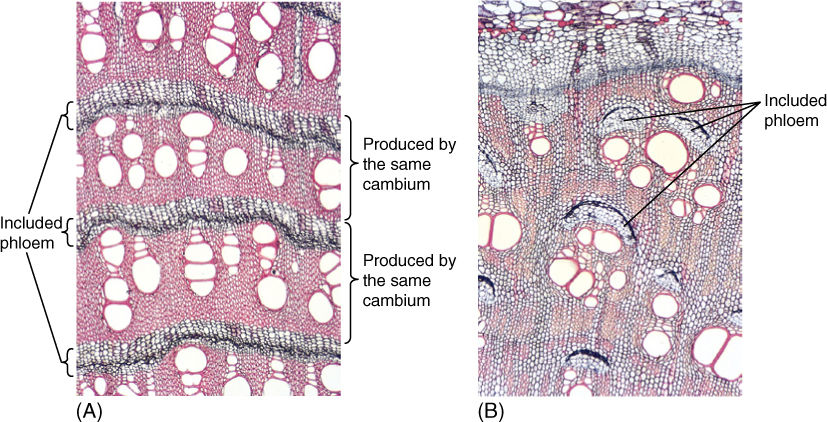
FIGURE 8-30 (A) This stem of Iresine shows bands of secondary xylem alternating with bands of included phloem. Each band of xylem and the phloem just external to it were produced by one cambium; the next set was produced by a different cambium (×40). (B) Bougainvillea also has included phloem, but it occurs in patches, not complete bands (×40).
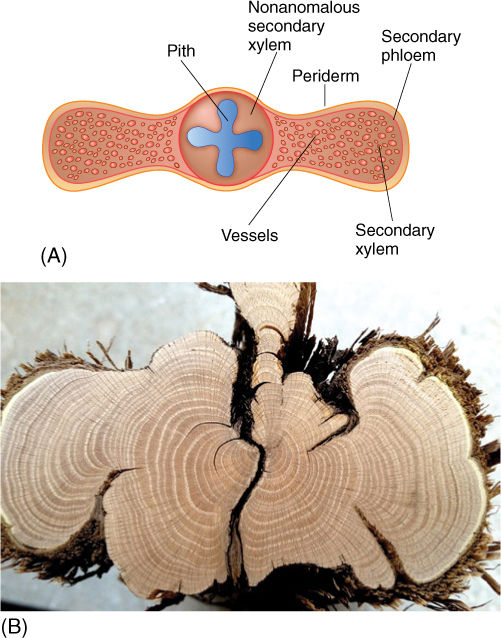
FIGURE 8-31 (A) The cambium on two opposite sides of this Bauhinia is rapidly producing cells, many of which differentiate into wide vessel elements. The two alternate portions of cambium produce very few cells, and all mature without much expansion. (B) Strips of the vascular cambium of sagebrush (Artemisia tridentata) periodically die while surrounding regions of cambium remain alive. This results in deep crevices in the trunk and branches, and there are no complete growth rings after the first strip of cambium dies.
Secondary Growth in Monocots
None of the monocots has secondary growth like that in gymnosperms, basal angiosperms, and eudicots, but some do become tree-like and “woody,” such as Joshua trees, dragon trees, and palms. The first two groups undergo a process of anomalous secondary growth; the palms have an unusual type of primary growth.
In Joshua trees (some members of the genus Yucca; FIGURE 8-32) and dragon trees (in the genus Dracaena), a type of vascular cambium arises just outside the outermost vascular bundles (FIGURE 8-33). It originates from cortex cells in the same manner that the interfascicular vascular cambium arises in eudicots. This cambium, however, produces only parenchyma; conducting cells are completely absent. Columns of some of the parenchyma cells undergo rapid division and produce narrow cells that differentiate into secondary vascular bundles containing xylem and phloem. The outermost cells of each bundle develop into fibers with thick secondary walls. The parenchyma cells that do not divide like this form a secondary ground tissue, the arrangement of which is almost identical to that of primary tissues. They are “woody” because of the fibers, and they have more conducting capacity and greater strength each year, so branching is feasible.

FIGURE 8-32 Joshua tree (Yucca brevifolia) is an arborescent monocot in the lily family. Because it has secondary growth, even though of an unusual type, the ability of its trunk to conduct increases, and both branching and increased numbers of leaves are feasible without adventitious roots of the type necessary for screwpine.
Unusual Primary Growth
Palm trees are unusual in that their trunks do not taper at the tips like those of eudicot or conifer trees, and they do not branch. The palm trunk is all primary tissue consisting of vascular bundles distributed throughout ground tissue; each bundle contains only primary xylem and primary phloem, all derived from the shoot apical meristem (FIGURE 8-34). A vascular cambium never develops, and true wood and secondary phloem do not occur; the trunk does not grow radially. The trunk is hard and “woody” because each vascular bundle is enclosed in a sheath of strong, heavy fibers.
A palm seedling does not have a full set of leaves and a wide trunk. For the first few years of life, the palm trunk becomes wider and the number of leaves increases. This happens without secondary growth because during the seedling years, palms produce numerous adventitious roots from the base of the short stem. Each root adds extra vascular bundles, and the portion of stem above each new root can have that many more bundles than it does below the root. For example, if the stem has 100 bundles at one point and if just above this it produces five adventitious roots that each have eight bundles, then above these roots the stem can have 140 bundles. This increase in width and addition of adventitious roots in palms is called establishment growth, a form of primary growth. At some point, this process ceases. No new adventitious roots are established, and the conducting capacity is set for the lifetime of the plant. This same type of primary growth occurs in other monocots with extremely broad stems such as corms and bulbs.
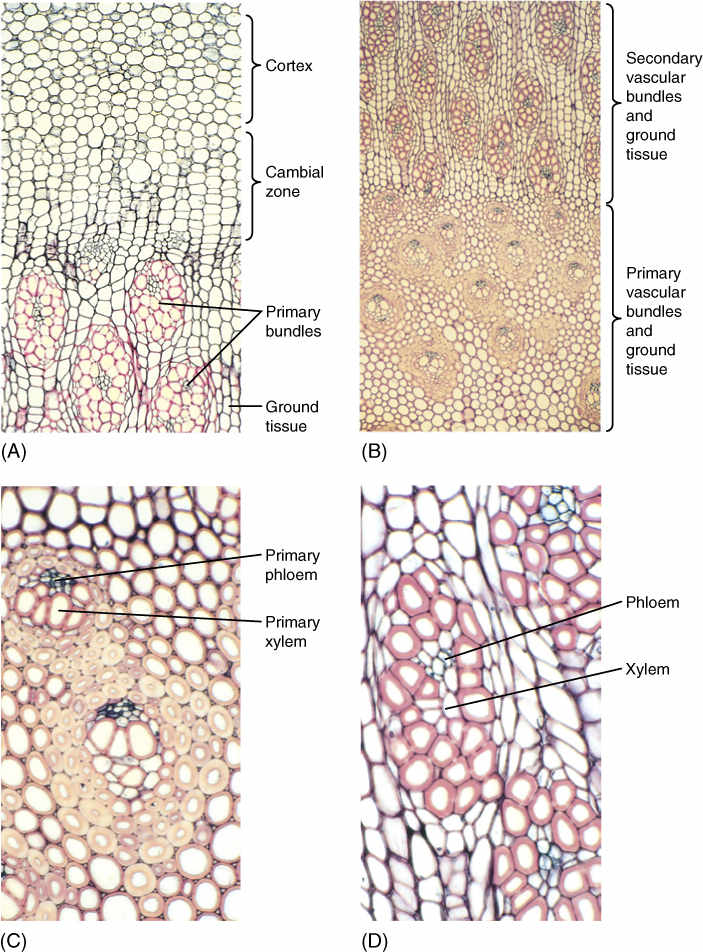
FIGURE 8-33 Secondary growth in arborescent monocots. (A) The broad zone of the vascular cambium region and immature secondary ground tissues are visible. The bundles are secondary, and a secondary cortex also forms (×50). (B) The inner part of the stem, with the primary vascular bundles and ground tissue on the bottom and the secondary bundles at the top (×15). (C) Primary bundles (×50). (D) Secondary bundles (×50).

FIGURE 8-34 Palm trees are monocots, so they have many vascular bundles rather than wood. This large palm trunk has hundreds of vascular bundles; the outermost, very dark bundles consist of just fibers, without xylem or phloem. This is shown about one-fourth life size.
Plants Do Things Differently
BOX 8-4 Thinking about the Growth of Wood
Although the growth of trees and their wood is perhaps the most complex type of growth you will ever try to understand, it really is not too difficult. Use your imagination to picture a complicated object in three dimensions and then imagine watching it grow. To start, imagine a tree without any leaves, flowers, or fruits. Now imagine removing the bark from all the woody areas, and then paint all the wood—every bit of the trunk, branches, and roots but not the twigs or root tips where there is epidermis. If you imagined this correctly, you have one single, continuous coat of paint that indicates where the vascular cambium is, and you have thousands of twig tips and root tips, each with an apical meristem. All active meristems of the tree are now visible in your imagination (the axillary buds and root primordia are inactive, but if your brain is up to it you can try to imagine those as well—just remember they are only on twigs and root tips, not in woody parts).
Hold that image in your mind, and now imagine you are making bread and have mixed yeast cells with water and flour and then kneaded them until the yeast cells are evenly distributed throughout the dough. Each yeast cell absorbs water and nutrients and then enlarges and divides; each daughter cell then repeats this process. All parts of the dough contain living cells and all are growing at the same rate. This is diffuse growth, and if we were doing this on the space station without gravity, our ball of dough would grow into a perfect sphere, becoming larger in all directions equally.
Plants have localized growth, and at the tip of every living twig and root there is a small mass of dividing cells, the shoot or root apical meristem. They have localized growth. If the meristems were to grow like the bread dough (equally in all directions), then the stems and roots would end up looking like lollipops. But all the meristem cells next to existing stem or root tissue act differently from the other meristem cells: They divide less and less and finally stop dividing, having become part of the new stem or root rather than continuing to be part of the meristem. If all meristem cells slowed their rate of division and became stem or root cells, the stem or root would stop growing, and they would not have indeterminate growth. It is important that some meristem cells become added to the growing stem or root, while at the same time some of the meristem cells continue to be meristem cells.
Fortunately, our own hair, nails, and skin have localized growth just like this. Let’s consider the growth of one of our hairs, as an example. Small blood capillaries at the base of the hair follicle bring water and nutrients to the base of the hair shaft, the area where hair cells are still alive and dividing. The hair “meristem” cells absorb the nutrients, grow, divide, and then do this again. Those dividing cells that are closest to the hair shaft (and farthest from the capillaries) gradually stop dividing and instead differentiate into dead hair cells. As time passes more hair meristem cells are added to the base of the shaft and our original cells are thus pushed upward. Each of our hairs grows at its base and is pushed upward out of the hair follicle; the outermost end of each hair is the oldest part (and thus the most likely to become a split end), and the base of the hair is the newest. Our hairs do not have growth rings like tree wood, but the outer end of each hair is old and the inner end is younger (think of the problems this causes for those who dye or bleach their hair).
When growth is localized instead of diffuse, the location of the meristem is important. In roots and stems the meristem is at the apex, the outermost part, so the apical meristem is pushed away from the plant body by the very shoot or root that it is producing. But the dividing cells of our hairs are located at the base of the hair; they are basal meristems and they remain in the same place; it is the hair itself that is forced to move as new cells are added to it. The same is true for our skin (the innermost basal layer contains the living, dividing cells, whereas the outer part that we see is dead) and nails (we only see the dead nail cells, and we can actually watch those cells being forced away from the fingernail base, the “quick,” by applying fingernail polish and seeing the new, fresh, dead unpainted nail cells being pushed out of the fingernail base).
Remember that one of the themes of this text is to think about alternatives and consequences. I am going to describe an alternative way of growing that does not exist; this is a theoretical alternative that has never evolved, as far as we know. We can imagine that some tree might have two vascular cambia located back to back and shaped like a hollow cylinder (the same shape as a real vascular cambium). The inner cambium would be a “wood cambium”: Its cells would divide, and those cells on the inner side closest to the existing wood would differentiate into new wood cells. We could say this wood cambium is an apical meristem because it is at the apex of the wood it is producing and it is being pushed outward as wood accumulates on its inner side. This wood cambium would be just like a shoot apical meristem except that it would be a cylinder instead of a tiny point, and, for a whole tree, it would have the shape of the paint we imagined in the first paragraph. Lying right to the outside of the wood cambium would be our hypothetical “phloem cambium”: All its cells would grow and divide, and the cells closest to the mature phloem would differentiate to be new phloem cells, and the rest of the phloem cambium cells would continue to be phloem cambium. Like the cells that produce our skin, nails, and hair, the meristem cells cannot be pushed deeper into the body; instead, the new tissues must be pushed out. So our phloem cambium would be a basal meristem and the new phloem would be pushed outward, just as our hair and nails are pushed outward. In this hypothetical, nonexistent model, our tree would grow wider because of its “apical” wood cambium and its “basal” phloem cambium lying back to back.
A minor problem is that as new cells are added to the wood, the wood becomes thicker and consequently so does its circumference: Its surface area increases. Because the “wood cambium” and the “phloem cambium” are on the surface of the wood, those two cambia also must become larger. We have a familiar example for understanding this: We have to be extremely careful around a newly born baby because the five boney plates that make up its skull are not yet solid; they are still growing at the edges (sutures) where each piece of bone meets its neighbors. As the baby’s brain grows, so does its skull, as the sutures between each plate add new cells to the edges of those plates. The bones in a baby’s skull become larger as a “meristem” of dividing cells along the edges of each plate adds new cells to each plate. And as each plate becomes larger, its perimeter becomes larger, so that “meristem” along the edges must increase, must become longer. It is just the same for the meristems that cover the surface of a growing cylinder. The baby’s skull adds enough new “meristem” cells so that all parts of the edge of each plate is completely covered in dividing cells and thus each plate grows everywhere along its edge.
Now let’s go back to the tree. Instead of having two meristems back to back, there is just one single layer of cells, just one meristem, the vascular cambium. As its cells divide, sometimes cells on the inner side stop dividing and differentiate as wood, and sometimes instead it is cells on the outer side that stop dividing and become phloem. In both situations it is critically important that some cells continue to divide and act as meristem cells. Because the addition of new cells to the wood causes the wood’s surface to increase in circumference and because the cambium is pressed tightly against the wood, the cambium’s circumference must also increase. The cambium has two options: Its cells could occasionally divide to make both daughter cells lie side by side within the cambium and neither one of those would turn into either wood or phloem. In this case the cambium itself would grow and have more cells. By doing this often enough the cambium will always have enough cells to completely cover the wood cylinder, no matter how broad it becomes and no matter how huge its circumference. Alternatively, rather than dividing to make new cambium cells, the cambium cells could just enlarge and become bigger cells. But as the tree continues to grow the cambial cells will also need to grow, and at some point they would reach a size that is just too large for a living, dividing cell to survive. At that point cell division would have to stop, and then the tree could no longer increase in width. This type of cambium did indeed evolve in one group of plants (species of Sigillaria), and the trunks it produced never grew to be very wide.
To summarize, shoots and roots of woody plants become longer as new cells are added to their apices in a form of localized, apical growth. Woody shoots and roots become wider through the action of a single vascular cambium that is only one layer of cells thick, and that layer produces xylem to its inner side and phloem to its outer side and it also produces new cambial cells. Each year, as the trunk or branch or root becomes longer, a new section of vascular cambium must differentiate, and it attaches itself to the preexisting cambium, and in the following year this will be repeated. At any given time the tree has only one cambium.
When trying to understand how trees grow, it is important not to make too much comparison with our bones. We refer to wood as a tree’s “skeleton” or that wood supports a tree the way our bones support and strengthen our body. But keep in mind that our bones are composed of living cells, most of which are encased in a layer of calcium phosphate. This may sound like a wood cell with a lignified secondary wall, but bone cells are alive and can remodel themselves, and bones are constantly taking some cells from certain spots, adding other cells to other spots, and remodeling the calcium (especially in women). Broken bones heal if set properly because our bodies can readjust the cells and the calcium phosphate, but the wood in a broken branch never heals. Some new wood may be laid down around it if the cambium survives, but the broken splinters inside never heal. And do not compare our growth in height with that of a tree; as we grow taller, the ends of our bones are constantly being reshaped and remodeled. New cells are added to the ends, and old cells are removed from other areas. This never, ever happens in plants; new wood cells are never added to the top end or bottom end of an existing piece of wood; instead, a piece of wood only grows wider through its vascular cambium. A branch or trunk becomes longer only because an apical meristem makes a new section of twig, and at some point that develops a new section of cambium that then starts to make wood, and this new cambium is attached to the end of the old cambium.
 At the Next Level
At the Next Level
1. Wood ecology. Wood of many different types has evolved by alterations in the diameter and number of vessels, thickness and lignification of fiber walls, presence or absence of tracheids, and modifications of perforation plates. Each of these factors affects the way a plant is adapted to a particular ecology. I recommend the book Ecological Strategies of Xylem Evolution by S. Carlquist (1975, University of California Press).
2. The beauty of wood. Many types of wood are used by artists to create beautiful objects such as bowls, trays, boxes, and furniture. An Internet search will produce many interesting articles. I recommend the book Identifying Wood: Accurate Results with Simple Tools by R. Bruce Hoadley (1990, Taunton Press) as an excellent introduction into the ways a person identifies unknown samples of wood; it also describes many types of artisanal wood. A second recommendation is Atlas of Woody Plants Stems: Evolution, Structure, and Environmental Modifications by F. H. Schweingruber, A. Börner, and E.-D. Schulze (2011, Springer Verlag), which has hundreds of magnificent photographs and light micrographs of wood structure and the plants that produce the wood. It is superb. Almost any arts and crafts fair has people selling items constructed from beautiful woods. Give some of these a close look and see if you can identify the woods; if not, ask the craftsman.
3. Wood as a source of lumber and chemicals. Wood is extremely useful to us as a source of lumber, paper, and chemicals such as resin and turpentine. Wood is essential to the sound of musical instruments such as pianos and violins. Wood can be sawn, chipped, ground to sawdust, glued, bent, and treated in many other ways. Some of these processes are modern, others are ancient, all will give you a deeper understanding of wood. In almost all cases, the anatomy of a piece of wood determines whether it will be suitable or useless for a particular product.
SUMMARY
1. Cells and tissues produced by apical meristems are primary tissues, whereas those produced by the vascular cambium and cork cambium are secondary tissues.
2. The vascular cambium contains fusiform initials that produce the elongate cells of secondary xylem and phloem, and ray initials that produce ray cells.
3. Secondary xylem contains the same types of cells as primary xylem, but the arrangement differs. Growth rings with early wood and late wood are usually present.
4. In an old trunk, branch, or root, the central xylem is heartwood; it is dry and its parenchyma has died. The outer, younger xylem is sapwood; it is involved in water conduction and its parenchyma is alive.
5. Cork cambium produces resistant cork cells; the cork and all exterior tissues are the protective outer bark. Lenticels permit oxygen diffusion into the organ.
6. Palms appear woody because their numerous vascular bundles contain many fibers. Certain other monocots have anomalous secondary growth, in which a vascular cambium produces secondary vascular bundles.
IMPORTANT TERMS
annual ring
anticlinal walls
axial system
bark
cork cambium
cork cell
diffuse porous
early wood
establishment growth
fusiform initials
growth ring
hardwoods
heartwood
included phloem
late wood
lenticels
nonstoried cambium
periclinal wall
periderm
phellem cell
phellogen
primary tissues
radial system
ray initials
rays
ray tracheids
reaction wood
ring porous
sapwood
secondary phloem
secondary tissues
secondary vascular bundles
secondary xylem
softwoods
storied cambium
tylosis
vascular cambium
wood
REVIEW QUESTIONS
1. Growth by apical meristems has been previously described. In woody species, additional tissues are produced in the stem and root by two other meristems, the _______________________________________ and the ________________________. The new tissues themselves are _____________________ (________________________) and ________________________ (________________________ and ________________________). They are the secondary tissues, and they constitute the plant’s secondary body.
2. Name at least three examples of woody plants that are trees and three that are shrubs.
3. Many herbs live for several years. How do they respond to the bottleneck of the limited conducting capacity of the first-year stem?
4. Which groups of plants have secondary growth? Which never do?
5. Imagine a tree that has a radius of 20 cm and that produces a new layer of wood 0.5 cm thick (the outer radius of the new wood is 20.5 cm and the inner radius is 20 cm). What is the cross-sectional area of the new layer of wood (CSA = pr2)? If each leaf needs 0.1 cm2 of wood to supply it with water, how many leaves can the tree have? If the tree produces a new ring of wood next year that is again 0.5 cm thick, how many leaves can the tree have next year (assume that wood conducts water for only 1 year, not 2 years)?
6. Woody plants are almost always perennial plants, often living for many years. Describe some advantages of this with regard to a plant’s ability to occupy a favorable site. Describe some disadvantages with regard to how long a plant must survive pathogens and harsh conditions.
7. It must be difficult for secondary growth to arise by evolution. How many times has it evolved in the 420 million years that vascular plants have been in existence? How many of those have become extinct?
8. What is the name of the vascular cambium that arises within vascular bundles? Between vascular bundles? After the two have formed, what is the shape of the vascular cambium? Is it a series of strips that are aligned up and down the trunk or root, or is it a complete cylinder?
9. Imagine a tree that is growing in springtime. All woody parts that were present last year have a vascular cambium, but all of the twigs that are brand new have just primary growth with vascular bundles. When will a vascular cambium form in these twigs? Will it connect to the old, pre-existing vascular cambium, or will it remain separate? Describe how this occurs.
10. Figure 8-3 is complex but easy to understand. Figure 8-3B shows a tree trunk, and this year is its fourth year of growth. A transverse section at the base of Figure 8-3B would show how many layers (growth rings) of wood (circle one: 1, 2, 3, 4)? Would a transverse section through the top of Figure 8-3B show any layer of wood this year? Would it show a layer of wood at the end of next year? Would there be any transverse section that does not show the pith and primary xylem; that is, are pith and primary xylem ever lost or destroyed by the formation of wood? The orange layer is labeled “Provascular tissue” in the top right of Figure 8-3B, but what is it labeled in the middle and lower right diagrams?
11. What are the two types of cells in the vascular cambium? Can either type be converted into the other?
12. Look at Figure 8-6. Part (A) shows the lower half of a fusiform initial (actually most fusiform initials would be much taller than this), and part (B) shows the same cell after it has divided and one of the daughter cells is developing into a xylem cell. Did the fusiform initial divide with a periclinal wall or an anticlinal wall as it went from part a to part b? Part (D) shows that the same fusiform initial has now produced a second cell that is developing into a phloem cell. Was that phloem cell produced by a periclinal or an anticlinal division of the fusiform initial? Part (F) shows the fusiform initial dividing and forming a second fusiform initial. Is this division occurring by a periclinal wall or an anticlinal wall?
13. Are fusiform initials parenchyma cells, collenchyma cells, or sclerenchyma cells? Do they have chloroplasts, chromoplasts, or proplastids?
14. Look at Figure 8-8. Vascular cambium is not present; it was located far above the top of the micrograph, but you can see there are six rows of wood cells. How many fusiform initials were involved in producing the wood visible in this figure? Look at all the cells in any particular row. Were all the cells of that row made by divisions of one fusiform initial or by many different fusiform initials?
15. Are ray initials longer or shorter than fusiform initials? Can they undergo both periclinal and anticlinal divisions or just one or the other?
16. Ray initials are typically grouped together in short vertical rows only one cell wide (________________________), two cells wide (________________________) or many cells wide (________________________).
17. True or false: Typically a vascular cambium never has large regions of only fusiform initials or ray initials.
18. What fraction of cells formed to the interior of the vascular cambium develop into secondary xylem, known as wood—less than half, exactly half, or more than half all cells?
19. In gymnosperms (conifers like Christmas trees), what types of cells occur in the axial system (the cells produced by fusiform initials)? Which cells are very rare (Hint: Table 8-2)? Wood of conifers is called ________________________ because a certain type of cell is absent from their axial system. Which type of cell is that?
20. In basal angiosperms and eudicots, what types of cells occur in the axial system? What is the only type of cell present in the rays of eudicots? The wood of eudicots is called ________________________ because it has the type of cell lacking in conifers. Which type of cell is that?
21. What types of cells are derived from fusiform initials? What types of cells are derived from ray initials? Is it theoretically possible to have a vascular cambium without ray initials or without fusiform initials?
22. Look at the woods of Figure 8-13. In part (B), all vessels have about the same diameter and are narrow, but the wood in part (C) has some very wide vessels (in the early wood) and some narrow ones (in the late wood, both types produced in the same year). The answer is not in the text, but would you think that these two species have different water conducting needs? Is it possible that one species lives in areas with very rainy spring times and the other lives in an area that is rather dry all the time?
23. What is a growth ring? How does early wood differ from late wood? Do all species of wood show strong differences between these two phases of a growth ring?
24. Why do some people prefer the term “growth ring” rather than “annual ring”? What can sometimes happen if a summer is unusually cool? Do you think this occurs more frequently in California or in Alaska?
25. If especially wide vessels are produced early in the growing season and only narrow, sparse vessels are produced later, the wood is said to be ________________________ porous; however, if vessels have similar size and abundance throughout a growth ring, the wood is ________________________ porous. Name three examples of each type.
26. What changes occur as sapwood is converted to heartwood?
27. What is a tylosis? How does formation of tyloses slow the spread of fungi in wood? Why doesn’t a tree make tyloses in vessels that are still conducting water?
28. The thickness of sapwood is some indication of how long a growth ring of wood is able to function. Look at Table 8-3. About how long does wood of catalpa function? How about wood of wild black cherry or honey locust?
29. Because secondary phloem is formed from the vascular cambium just as secondary xylem is, it too has an ________________________ system and a ________________________ system. Why do the size, shape, and number of phloem rays match those of xylem rays?
30. In a cross-section of a tree, where are the oldest growth rings—in the outer region or nearer the pith? Where is the oldest secondary phloem—near the outside of the tree or near the cambium?
31. What causes the outermost tissues of a woody stem or root to become pushed outward and expanded?
32. The layer of cells that produces cork has a technical name and an ordinary name. Give each name.
33. Periderm consists of at least two types of cells. Sometimes a third is present. What are these three types of cell?
34. Why does a layer of periderm offer only temporary protection? When the plant makes a new layer of cork cambium, does it make the new layer to the outer side of the failing periderm or to the inner side of it, deeper in the secondary phloem?
35. In which tissues does the first cork cambium form? When does it usually arise? In which tissues do later cork cambia form?
36. Are geraniums herbs or woody plants? Do they ever form bark (Hint: Figure 8-23)?
37. What type of plant produces the cork used for wine bottles and cork boards—pines, oaks, maples, eucalyptus, or none of these (Hint: Figure 8-25)?
38. What is the function of lenticels? How are intercellular spaces important for this function?
39. Do roots form wood and bark, or are these secondary tissues present only in stems?
40. Describe the anomalous secondary growth in roots of sweet potatoes. What is the selective advantage of this unusual type of secondary growth?
41. Describe the formation of included phloem. How did the included phloem of Iresine in Figure 8-30A become surrounded by xylem?
42. Describe the secondary growth of monocots like Joshua trees and dragon trees. What are secondary vascular bundles?
43. Describe the growth of a palm seedling. For the first few years of life, the seedling becomes wider, but without secondary growth, how is it able to do this? Each adventitious root adds something. What is it? If the palm stem has 100 vascular bundles and then forms five new adventitious roots with eight bundles each, how many vascular bundles can the stem have above these new adventitious roots? Would the stem be wider or narrower above these adventitious roots?
44. Why can a monocot like an iris branch and increase its number of leaves? Is the fact that the shoot is a rhizome with adventitious roots important? Is water transported from one end of the shoot to the other?
45. What is the name of the analysis of tree rings? How is that used to study past climates? How is it used to establish the date when ancient buildings and ships were constructed?
46. In tree ring analysis, what is a floating sequence? Why has the sequence for the Middle East been floating, whereas we know the exact dates of each ring in the sequences for North America and Europe? What is so important about the volcanic eruption in 1628 BCE? Do you think it is possible that the unusual ring in North American tree sequences was produced by a volcanic eruption in North America and the unusual ring in the Middle East sequence was caused by a different phenomenon that occurred in a different year? If so, then has the floating sequence been anchored?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art