Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Genetics and Evolution
Algae and the Origin of Eukaryotic Cells

Chapter Opener Image: What does a whale have to do with algae? Whales eat microscopic animals called krill, and krill eat algae. Without algae, there would be no whales, and for that matter, there would be no other marine animals: Algae are the basis of the food chain in the oceans, as well as in most rivers and lakes. The bodies of algae are organized differently than those of plants, so algae are considered to be close relatives of plants but not actually plants. Like plants, almost all algae are photosynthetic (there are parasitic algae) and they produce a significant amount of the oxygen we breathe. Algae are the ultimate food of all marine animals. Many algae are unicellular or have very simple multicellular bodies that have not changed much in millions of years, so they are often considered to be “primitive.” But true plants have existed for more that 420 million years, and none has ever become as well adapted to living in the oceans as algae are.
OUTLINE
✵ Concepts
✵ Origin of Eukaryotic Cells
- DNA Structure
- Nuclear Structure and Division
- Organelles
- Origin of Mitochondria and Plastids: The Endosymbiont Theory
✵ Characteristics of Various Groups of Algae
✵ Green Algae
- Body Construction in Green Algae
- Life Cycles of Green Algae
- Representative Genera of Green Algae
- Green Algae and Embryophytes
✵ Red Algae
✵ Brown Algae and Their Relatives: The Heterokonts
- Brown Algae
- Diatoms
- Yellow-Green Algae
- Golden-Brown Algae
✵ Dinoflagellates
- Oomycetes
✵ Euglenoids
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Summarize the basic steps in the evolution of algae.
✵ Name three differences between prokaryotes and eukaryotes.
✵ Explain the endosymbiont theory.
✵ Describe how various algae evolved by means of primary endosymbiosis and secondary endosymbiosis.
✵ List three significant attributes of green algae.
✵ Recall six evolutionary possibilities from a motile single cell.
✵ Summarize meiosis and syngamy in relation to green alga reproduction.
✵ Give an example of each of the following types of algae: unicellular, motile colonial, filamentous, laminar, coenocytic, and parenchymatous.
✵ Recall what sets red algae apart from other algae.
✵ List three characteristics of brown algae.
✵ Compare brown algae to green organisms.
✵ Describe the morphology of a diatom.
✵ Provide three unique characteristics of dinoflagellates.
 Did You Know?
Did You Know?
✵ Many algae in lakes and streams are mistakenly called moss.
✵ Many algae live in soil, more live in freshwater, and many inhabit oceans, but none lives at water depths where sunlight does not penetrate.
✵ Algae are often referred to by their color, such as green algae, red algae, brown algae, and so on; these differ not only in their pigments but also in basic anatomy and reproduction.
![]() Concepts
Concepts
Life originated over three billion years ago as simple prokaryotic organisms (FIGURE 19-1 and see the timeline on the inside of the back cover). Some time later, perhaps less than a billion years after that, eukaryotic cells had come into existence. At first, these too were extremely simple, probably having the rudiments of a membrane-bounded nucleus, endoplasmic reticulum, microtubules, and so on, but they did not have mitochondria or plastids, at least not initially. Mitochondria arose first, as an early eukaryotic cell engulfed but did not digest a bacterium capable of aerobic respiration. The two organisms lived together, one inside the other, and both benefited: This is an endosymbiosis. Over hundreds of millions of years, this gradually evolved into a more sophisticated form of eukaryotic cell with real mitochondria, and as it did so, it diverged into many species. Some of these species died out rather quickly; others were more successful and persisted, also diversifying into many forms. Currently, there are numerous groups of organisms, often called protozoans and algae (FIGURE 19-2) that are the living descendants of clades that originated from those early steps in eukaryote evolution. Eventually, one of those would establish a clade that much later diverged into animals and fungi—multicellular organisms that have nuclei and mitochondria but not plastids (FIGURE 19-3).
Another line of early eukaryotes, ones that did have mitochondria, entered another endosymbiosis, this time with a photosynthetic cyanobacterium, which later evolved into a chloroplast. This line gave rise to algae, including green algae which in turn produced true plants—embryophytes. As you can imagine, numerous modifications are needed to convert an undigested cyanobacterium into the chloroplasts in angiosperm leaves. Fortunately, during this long process, several clades originated that still have some extant members whose plastids have numerous prokaryote characters; chloroplasts of red algae especially resemble cyanobacteria.
We are fortunate that many organisms still exist that have many ancestral characters, characters that we believe were present in early eukaryotes. These are almost always unicellular and in the past were grouped together in a kingdom called “Protista,” but this was a grade level of classification: They were grouped together because they were considered to represent a low grade, a low level of evolution, they lacked derived characters of plants, animals, and fungi. We no longer group organisms on the basis of what they lack; therefore, “Protista,” the formal name, has become “protista,” the informal name. More importantly, however, DNA sequencing studies, corroborated by electron microscopy and other techniques, are revealing the phylogenetic relationships of these organisms (Figure 19-3).
This chapter focuses on the origin and diversification of photosynthetic eukaryotic cells. This includes the various types of algae, especially the green algae, which are closely related to the ancestors of true plants, the embryophytes. Reproductive structures are the critical factors in distinguishing algae from plants. Algae are defined as photosynthetic individuals whose reproductive structures are completely converted to spores or gametes that, when released, leave nothing but empty walls (FIGURE 19-4). In plants, the reproductive structures are always complex and multicellular, and only some of the inner cells become reproductive. Outer cells constitute a protective layer and persist after the reproductive cells are released. This may seem trivial, but the algal method of reproduction is a cellular process, reflecting the simple, cellular level of organization most species have throughout their bodies. Reproductive structures of plants involve two tissues—reproductive and sterile—functioning together as an organ; this reflects a more complex organ level of integration in the entire body.
![]() Origin of Eukaryotic Cells
Origin of Eukaryotic Cells
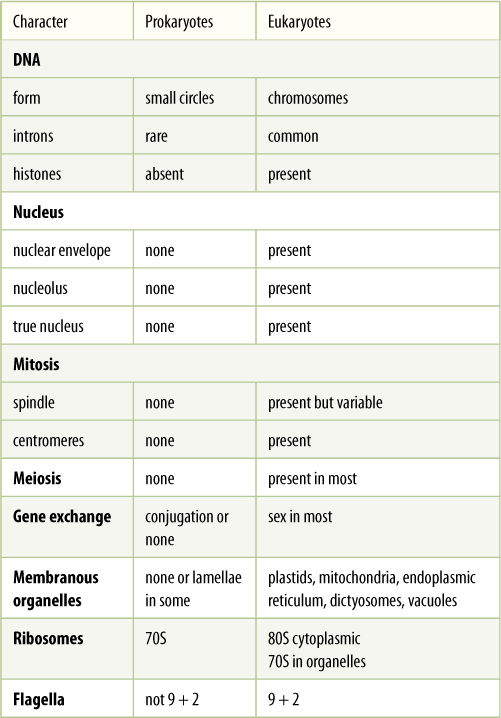
The differences between prokaryotes and eukaryotes are deep seated, involving many of the most basic metabolic processes and cellular organization (TABLE 19-1).
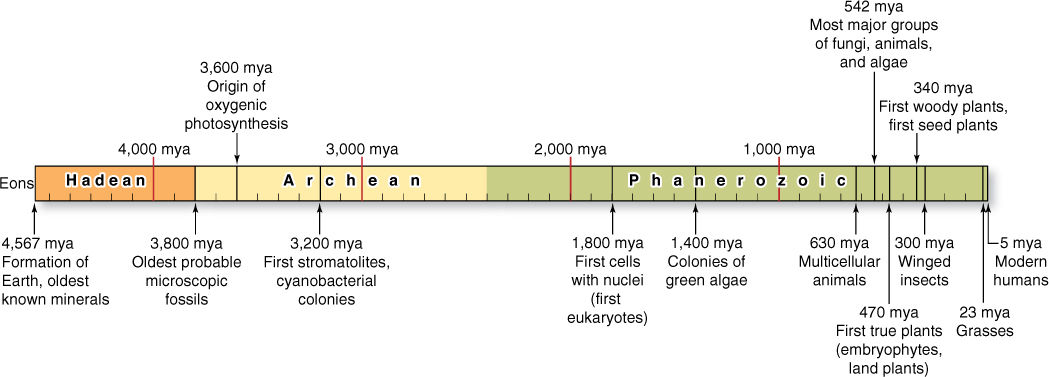
FIGURE 19-1 The Earth’s history has been divided into three eons by geologists: the Hadean Eon, the Archaean Eon, and the Phanerozoic Eon. Significant events are indicated. (mya = million years ago)
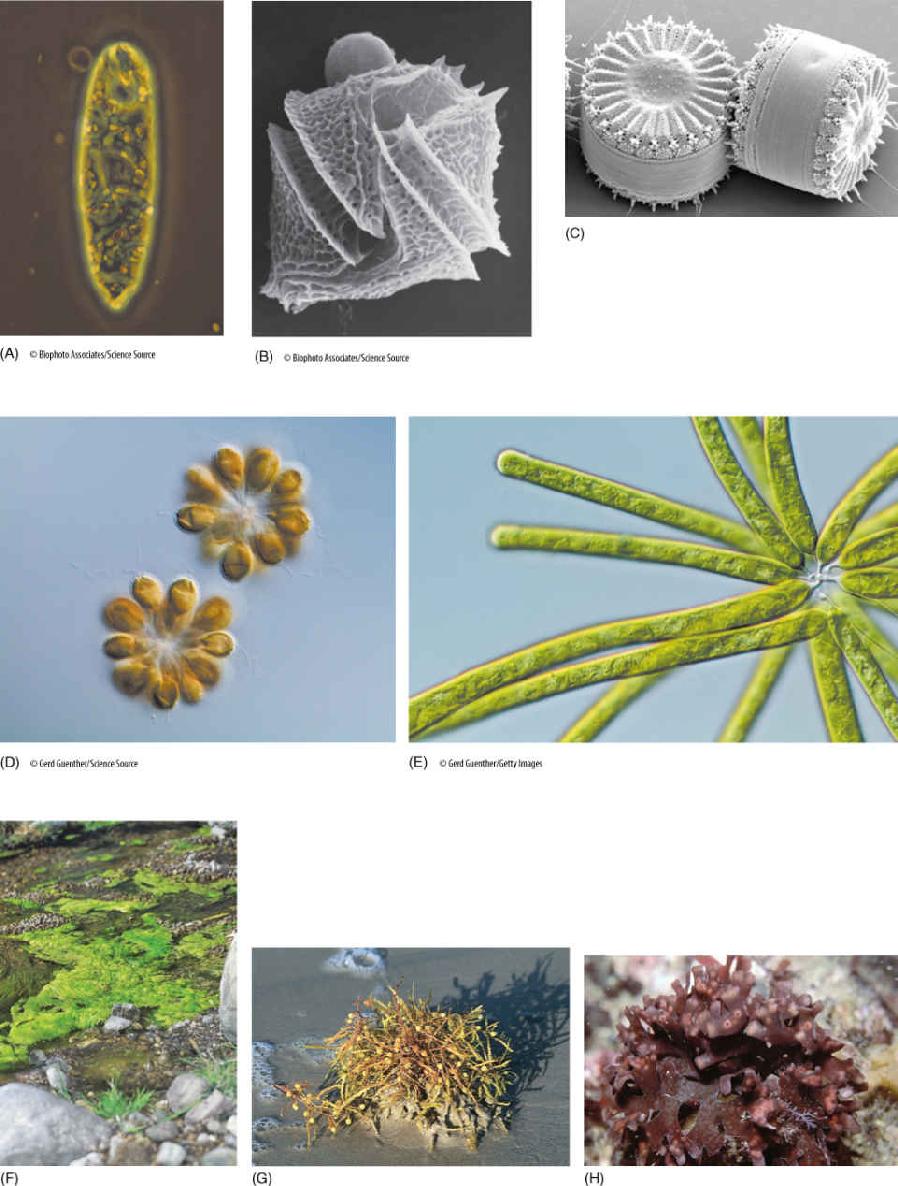
FIGURE 19-2 (A) Euglenoid algae. (B) A dinoflagellate. (C) Centric diatoms. (D) Golden-brown algae. (E) Yellow-green algae. (F) Green algae. (G) Brown algae. (H) Red algae.
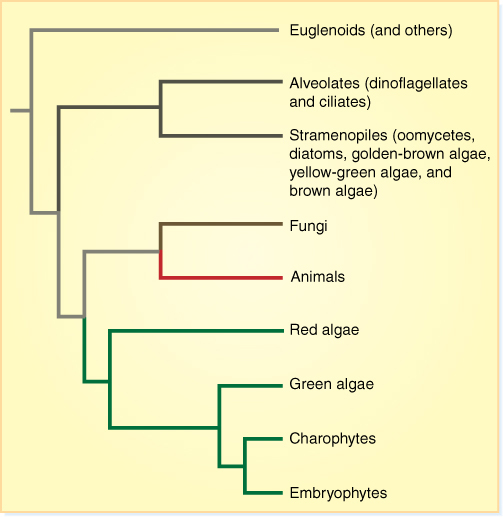
FIGURE 19-3 One recently proposed cladogram of the phylogeny of algae. The organisms that we call algae are parts of numerous clades, but they do not make up one single monophyletic clade. The ancestor that is common to all algae is also common to animals, fungi, and plants: It is the ancestor of all eukaryotes. Consequently, we use “algae” as an informal name.
DNA Structure
In prokaryotes, DNA is “naked,” not complexed with proteins, its numerous negative charges being neutralized by calcium ions instead. Each genome is just a short circle of DNA, often containing only approximately 3,000 genes. Prokaryote DNA contains few introns, and mRNA processing is relatively simple.
In eukaryotes, DNA is more elaborately packaged, being complexed with nucleosome histones and forming chromosomes. Each DNA molecule is long, capable of carrying thousands of genes. Nevertheless, no genome ever consists of just a single DNA molecule; at least two occur per haploid set, and 10 to 20 are more common. Many eukaryotic genes contain introns, and large segments of DNA do not code for any type of RNA.
TABLE 19-1 Differences Between Prokaryotes and Eukaryotes
Nuclear Structure and Division
The DNA of prokaryotes lies directly in the cytoplasm; no nucleus is present. As a prokaryotic cell grows, its plasma membrane expands; the DNA circles attached to it are separated from each other by cell growth. The cell then pinches in two.
The nuclei of plants are virtually identical to those of animals and fungi in structure, metabolism, mitosis, and meiosis. Apparently these three clades diverged only after eukaryotes and their nuclei had become quite sophisticated; however, nuclei are complex organelles, and mitosis and meiosis are intricate processes with many steps. Certainly plant and animal nuclei could not have arisen from prokaryotic nucleoids quickly; many intermediate steps spanning hundreds of millions of years must have been involved.
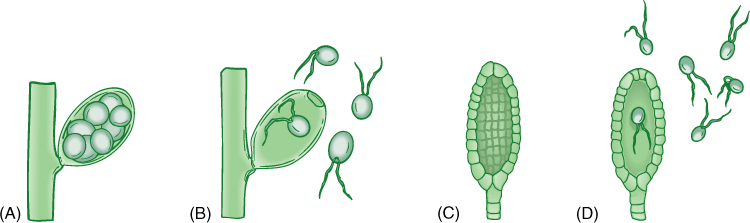
FIGURE 19-4 Algae are distinguished from plants (embryophytes) primarily by the nature of the reproductive structures. (A) and (B) In algae, the sporangium or gametangium may be either one large cell, as shown here, or multicellular, but every cell is converted into a reproductive cell. When reproductive cells are released (B), only the wall of the original cell is left. (C) and (D) Sporangia and gametangia of plants are always multicellular and only the inner cells differentiate into spores or gametes; after they are liberated (D), a residual layer of sterile cells remains.
Most of the DNA of eukaryotes is located within a nucleus, separated from the cytoplasm by two nuclear membranes with nuclear pores; this is the source of their name (eukaryote = true nucleus). A nucleolus is present as well. Eukaryotic nuclei are typically haploid or diploid, having only one or two copies of each of their various chromosomes, and thus, the rather haphazard prokaryotic method of simple membrane expansion would not work reliably. Instead, mitosis ensures that each progeny cell receives one of each type of chromosome. In almost all eukaryotes, meiotic nuclear division occurs as part of sexual reproduction. The critical phenomena are pairing of maternal and paternal homologous chromosomes followed by crossing-over and genetic recombination. These complex procedures result in an interchange of alleles, such that a great diversity of spores or gametes is produced.
Several groups of organisms with unusual nuclear characteristics may represent lines of evolution that originated earlier in the history of eukaryotes than did plants and animals. Some have both an intranuclear and an extranuclear spindle, and in some, the nuclear envelope and nucleolus either do not break down at all or do so only late in mitosis. Gaps may form in the nuclear envelope, and bundles of microtubules pass completely through the nucleus.
Nuclei of dinoflagellates have no histones. They do have what appear to be chromosomes, but these remain condensed at all times, undergoing only a slight uncoiling during interphase.
Organelles
Prokaryotes lack membrane-bounded organelles: Their cytoplasm is rather homogeneous, containing only ribosomes and storage granules. A few species have gas vacuoles, and photosynthetic species have folded membranes that project into the cell. All eukaryotes have nuclei and mitochondria, and plants and algae also have plastids. In addition, dictyosomes, endoplasmic reticulum, vacuoles, and vesicles are components of the eukaryotic endomembrane system. These membranes result in the division of the cell into numerous compartments, permitting several types of highly specialized metabolism to occur within the same cell. Ribosomes of prokaryotes are 70S, being smaller and denser than the 80S ribosomes in the cytoplasm of eukaryotes. Plastids and mitochondria also have 70S ribosomes instead of 80S ribosomes.
Flagella and cilia are remarkably uniform in eukaryotes, consisting of a 9 + 2 arrangement of microtubules. Only a few prokaryotes have flagella, and these have a totally different type of construction, never a 9 + 2 arrangement. They are not composed of microtubules or tubulin.
Origin of Mitochondria and Plastids: The Endosymbiont Theory
Prokaryotes represent the types of organisms that arose first. Until the early 1970s, it was assumed that some had given rise to eukaryotes by gradually becoming more complex and developing an endomembrane system, parts of which eventually specialized so much they became mitochondria and plastids. This was the autogenous theory.
In the 1960s, the endosymbiont theory was revived. In 1905, K. C. Mereschkowsky had speculated that plastids and mitochondria might be prokaryotes living inside eukaryotic cells. At that time, there was no way to test the concept, but by the late 1960s, plastids and mitochondria were discovered to have their own DNA and ribosomes, both having prokaryotic features. Furthermore, these organelles divide by a type of cleavage similar to that of prokaryotes, and they also lack microtubules. Before long, other prokaryotic traits were discovered: Their DNA is a small closed circle of naked DNA, and organelle genes are organized like prokaryotic genes. Their ribosomes are sensitive to the same antibiotics that interfere with prokaryotic ribosomes.
Origin of Mitochondria
The endosymbiont theory postulates that a prokaryotic cell had evolved to the point of having some eukaryotic features such as 80S ribosomes and a nuclear envelope (FIGURE 19-5). It would have been heterotrophic, living by engulfing and digesting other cells. One of these early eukaryotic cells engulfed a type of bacterium but then did not digest it, perhaps because the larger cell had inefficient digestive enzymes or because the engulfed cell had a more resistant membrane. Such an association might be advantageous to both: The engulfed cell could be efficient at aerobic respiration but poor at obtaining nutrients. If it could obtain sugars from the larger cell and “leak” ATP back to it, both cells might be better off. Each species might have survived better when in an endosymbiosis than when each partner was living independently. Such an endosymbiosis would still have many obstacles to overcome—increasing the compatibility between the two cells and coordinating growth, development, and reproduction—but the descendants of the engulfed organism had the potential to evolve into a mitochondrion. DNA sequence data indicate that mitochondria are most closely related to bacteria known as proteobacteria and that the endosymbiosis occurred approximately two billion years ago. All known extant eukaryotes have mitochondria—no living eukaryote is primitively amitochondrial.
Origin of Plastids
Plastids could arise similarly if the engulfed partner were photosynthetic (Figure 19-5). Because all chloroplasts contain chlorophyll a and produce oxygen but none has bacteriochlorophyll, cyanobacteria were suspected to be the ancestors. Two groups called the green and purple bacteria are unlikely candidates because they have very different pigments and do not produce oxygen when they photosynthesize. Prochlorophytes are a type of cyanobacteria that arouse great interest because they have both chlorophyll a and b and some lack phycobilin pigments. The first prochlorophyte discovered, Prochloron, not only resembles chloroplasts biochemically but also exists as an obligate symbiont inside marine invertebrates, called didemnid ascidians, supplying them with carbohydrates whenever the ascidians lie in sunny areas. A second prochlorophyte, Prochlorothrix, has been discovered growing freely in lakes in Holland. 16S rRNA sequencing has now shown that chloroplasts, Prochlorothrix, and Prochloron are all definitely closely related, but neither cyanobacterium is the immediate ancestor of chloroplasts. Instead, all three share a common ancestor.
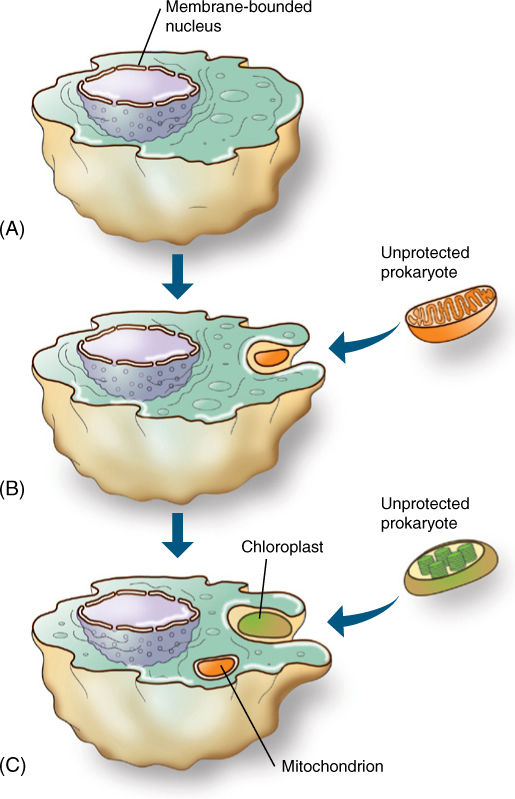
FIGURE 19-5 The endosymbiont theory postulates that a prokaryote (A) evolved to the level of having a membrane-bounded nucleus; this must have been an early step because all eukaryotes have a nuclear envelope and all are rather uniform in structure. (B) The early eukaryote engulfed but did not digest a prokaryote, which then evolved into a mitochondrion. (C) Finally, some descendants of that early eukaryote would engulf but not digest a prochlorophyte type of cyanobacterium, which would evolve into a plastid.
Chloroplasts have originated several times, but not in the way we had anticipated. Chloroplasts in green, red, brown, and other types of algae differ from each other in so many ways that we had expected that various types of cyanobacteria and early eukaryotes had formed protochloroplast endosymbioses. However, accumulating data from DNA sequences, electron microscopy, and biochemistry are consistent with a very different hypothesis: There was a single primary endosymbiosis and then multiple secondary endosymbioses. The primary endosymbiosis gave rise to a clade containing red algae, green algae, and a small group called glaucophytes (Figure 19-3). One feature of glaucophyte chloroplasts is that they still actually produce a thin film of cyanobacterial wall between themselves and the cell. Red algal chloroplasts have chlorophyll a (but not b) and the cyanobacterial pigment phycobilin, organized into particles called phycobilisomes (FIGURE 19-6). Green algal chloroplasts have no trace of bacterial wall or phycobilin, but instead have chlorophylls a and b and carotenoid accessory pigments, all of which are similar to chloroplasts in true plants (embryophytes). Glaucophytes are just a small group of algae. Red algae are a larger and more diverse group. Green algae are extremely numerous, and some are the sister group of embryophytes. Land plants are highly modified descendants of green algae that are in turn derived from the eukaryotes of the original primary chloroplast endosymbiosis.
Several events of secondary endosymbiosis produced other lines of algae. Euglenoids arose after an early eukaryote engulfed an entire green alga: A eukaryote engulfed another eukaryote. Since then, the cell of the green alga has become so reduced that mostly just the green algal chloroplast remains.
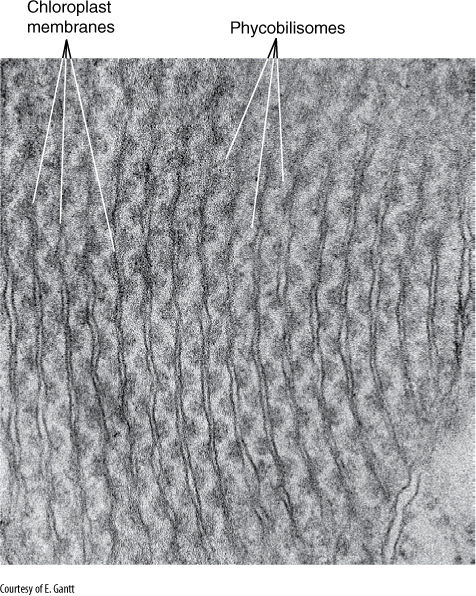
FIGURE 19-6 Internal membranes of red alga chloroplasts do not form grana stacks like those of green plants. Their phycobilisomes are visible, attached to the membrane. Porphyridium (×115,000).
Another group of early eukaryotes, called heterokonts (or stramenopiles), were involved in one or several endosymbioses with entire cells of red algae, and these gave rise to brown algae, golden-brown algae, yellow-green algae, and diatoms. Heterokonts are named for the fact that all have two flagella of different types: One flagellum is smooth; the other has hairs (FIGURE 19-7). All heterokonts have this flagellum heteromorphy; thus, they seem to form a monophyletic clade, but not all have chloroplasts. We think that heterokonts diversified somewhat, and then some entered into secondary endosymbioses and became photosynthetic, whereas others did not. A lack of chloroplasts in these heterokont protists is an ancestral condition. We are not yet certain whether there was more than one secondary endosymbiosis that produced pigmented heterokonts; there is a diversity of structure and biochemistry in the extant chloroplasts, as the red alga symbiont became modified in different ways in different lines (FIGURE 19-8). Most are similar in having chlorophylls a and c (not b and not phycobilins), and most have four chloroplast membranes, not just the two (outer and inner) of red algae, green algae, glaucophytes, and true plants. Of the four chloroplast membranes, the two innermost membranes correspond to the original inner and outer membranes of the red alga chloroplast, a third membrane that encompasses these and corresponds to the red alga plasma membrane, and an outermost fourth membrane that corresponds to part of the heterokont cell’s endoplasmic reticulum (called its cER for chloroplast ER; FIGURE 19-9). No heterokont chloroplast still has any remnant of the red alga cell wall, but some have a tiny bit of red alga nucleus, called a nucleomorph, which still has a nuclear envelope and a few genes. These cells have four types of DNA: heterokont eukaryotic nucleus, red alga eukaryotic nucleomorph, chloroplast prokaryotic DNA circles, and mitochondrion prokaryotic DNA circles.
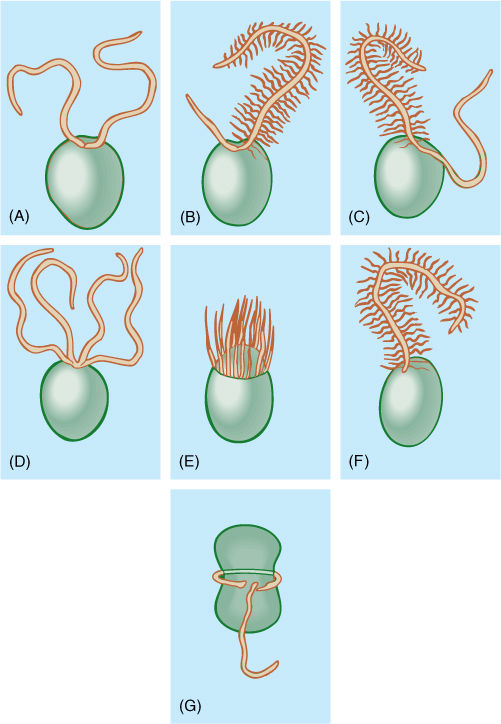
FIGURE 19-7 The most common arrangements of flagella in algae. (A) Two equal whiplash. (B) A short whiplash and a long tinsel. (C) Two equally long flagella, one of each type. (D) Four whiplash. (E) Numerous flagella located in a ring (rare). (F) One tinsel flagellum (mostly just in some brown algae). (G) One flagellum trailing, one in a transverse groove (dinoflagellates).
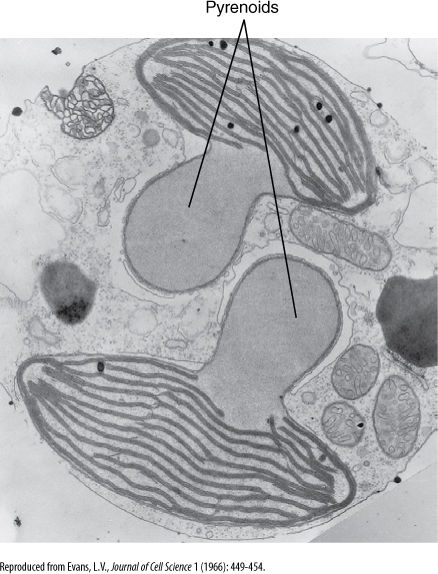
FIGURE 19-8 Brown alga chloroplast membranes associate into stacks of three or, less often, stacks of two. The large clear structures are pyrenoids, involved in polymerization of sugars into polysaccharides.
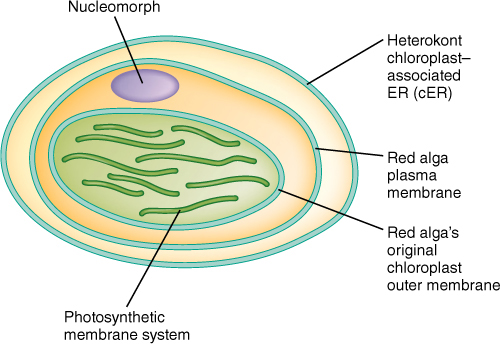
FIGURE 19-9 Many heterokont chloroplasts have four membranes: an outermost cER membrane, an inner membrane that corresponds to the plasma membrane of the red alga endosymbiont, and two innermost membranes corresponding to those of the chloroplast of the red alga. The innermost, photosynthetic membrane also corresponds to that of the original cyanobacterium. This entire structure would be located within the cytoplasm of a heterokont algal cell.
Types of Cytokinesis
Several types of cytokinesis occur in algae. Many groups of unicellular algae have no wall, their plasma membrane being their outermost surface. During cell division, the plasma membrane pinches in two, being pulled inward as a cleavage furrow, a process remarkably similar to cytokinesis in animals.
In almost all algae with walls, cell division is similar to that of plants. It involves a phragmoplast, consisting of short microtubules oriented parallel to the spindle microtubules. In some green algae, cytokinesis occurs by a phycoplast, with microtubules oriented parallel to the plane where the new wall will form, which is perpendicular to the orienta tion of the spindle. A phycoplast may be associated with division either by furrowing or by cell plate formation. Embryophytes arose from green algae that divide with a phragmoplast rather than a phycoplast.
![]() Characteristics of Various Groups of Algae
Characteristics of Various Groups of Algae
The descriptions of algae will begin with green algae because they are in the clade that encompasses not only the ancestors of embryophytes but also the primary endosymbiosis that originally produced chloroplasts. Red algae are described after that, and then attention is turned to the lines that resulted from the secondary endosymbiosis of plastids.
![]() Green Algae
Green Algae
From an evolutionary standpoint, the green algae (TABLE 19-2) constitute an extremely important group: Not only could some early green algae organize complex differentiated multicellular bodies, but some moved onto land—the ancestors of embryophytes, true plants. Perhaps because most of the animals and plants with which we are familiar are multicellular and terrestrial, we tend not to appreciate just how complex these phenomena are. Consider that organisms spent as many as 1 billion years at the level of unicellular organization (Figure 19-1); then among algae true multicellularity evolved only a few times. The transition to having a complex photosynthetic body capable of living on land was so difficult that it appears to have occurred only once.
Green algae have remarkable developmental and metabolic plasticity: They are resilient and survive many types of disturbances and changes. Mutations that cause cells to adhere after cytokinesis have not been lethal, nor were many mutations that affected swimming, orientation of cell division, or coordination of karyokinesis and cytokinesis. Bodies composed of several types of specialized cells have not evolved in most other algal groups. Although brown algae and red algae do show considerable sophistication in certain types of multicellular bodies, they are metabolically intolerant of ecological changes; few can live in fresh water, soil, air, or inside animals as many green algae do.
TABLE 19-2 Divisions of Algae

The diversity in green algae is tremendous. Examining it will give you a basis for understanding the diversity possible among living organisms and appreciating that the metabolisms and organizations of humans and flowering plants are by no means the only solutions to biological problems.
Body Construction in Green Algae
The most relictual and simple body is probably the motile single cell. Numerous green algae are unicellular. From this, many evolutionary possibilities exist:
1. Motile colonies (FIGURE 19-10A and Figure 19-18): If cells adhere loosely, the resulting structure is a colony, not an individual organism. All cells are similar, and none is particularly specialized; there may be some differentiation into two or three somewhat distinct cell types.
2. Nonmotile colonies occur if the cells lose their flagella or never develop them (FIGURE 19-10B). Although a nonmotile cell is simpler than one with flagella, the flagellated form is believed to be ancestral because all motile algae have the same type of flagella, the 9 + 2 arrangement of microtubules.
3. A filamentous body results if cells are held tightly by a middle lamella and if all cells divide transversely (FIGURE 19-11A and B and Figure 19-19). If occasional cells undergo longitudinal division, the filament branches. Often some portions of the body differ from others; for example, one end may serve as a means to attach the filament to a rock (a holdfast), or other parts may produce spores or gametes.
4. A membranous body results if the orientation of cell divisions is controlled precisely such that all new walls occur in only two planes (FIGURE 19-11C). The result is a sheet of cells that can become more extensive but remains thin. A membranous body is more strongly affected by currents and wave action and more likely to be torn than is a filamentous body.
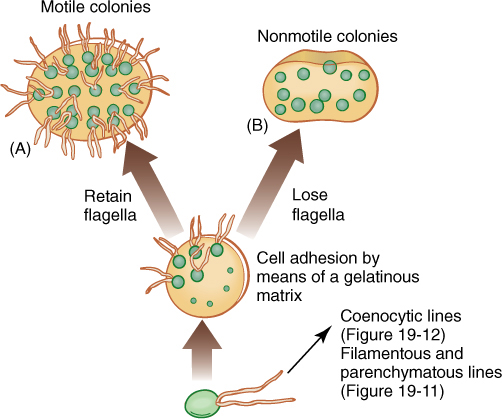
FIGURE 19-10 Evolution of colonial body type in green algae. The association of cells is controlled, not random. Depending on the species, each colony has a specific number of cells held in a particular shape such as a sphere, a flat plate, or a curved plate. No cell is free to divide at random.
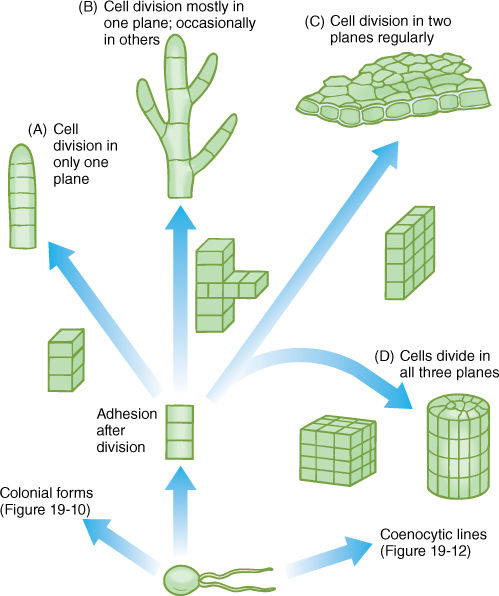
FIGURE 19-11 Evolution of filamentous, membranous, and parenchymatous body types in green algae. The formation of a multicellular body requires control over timing, position, and orientation of cell division, along with cell-cell adhesion mediated by a middle lamella. (A) All cells divide in only one plane, resulting in a uniseriate filament. (B) If cells occasionally divide in a different plane, the filament branches. (C) A two-dimensional sheet of cells results if all cells divide in two dimensions regularly. (D) A three-dimensional body is produced if cells divide regularly in all three planes. All cells share plasmodesmata with all adjacent cells; this is a true parenchyma tissue.
5. If cell division occurs regularly in all three planes, a bulky, three-dimensional parenchymatous body results (FIGURE 19-11D and Figure 19-22). All cells are interconnected by plasmodesmata, and a true parenchyma tissue is formed.
6. A coenocytic or siphonous body results if karyokinesis occurs without cytokinesis, and giant multinucleate cells result (FIGURE 19-12 and Figure 19-21). The cells can grow to several centimeters in diameter, but they usually remain fairly simple despite their size. Some coenocytes, such as Acetabularia, can be surprisingly complex (FIGURE 19-13).
Evolutionary development of control over the orientation of cell division and adhesion must have been difficult. The fossil record indicates that the ability to grow as a siphonous form came into existence 230 million years earlier than the ability to organize even the simplest filament.
Life Cycles of Green Algae
The angiosperm type of life cycle, an alternation of heteromorphic generations, can be traced to green algae, and the theory of life cycles can be discussed here. In unicellular algae that have no sexual reproduction, such as euglenoids, the life cycle is really just the cell cycle; mitosis and cytokinesis constitute reproduction. However, with the evolution of sex, two processes are critical: meiosis, which segregates out haploid sets of chromosomes, and syngamy, which brings two sets back together.
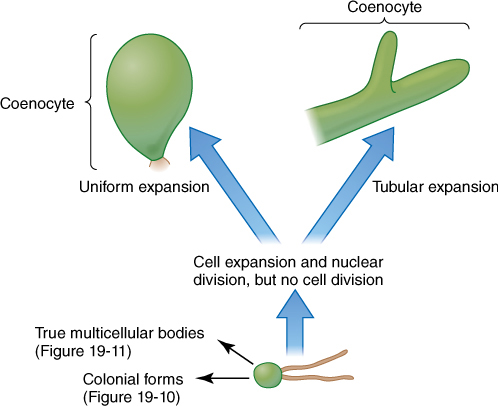
FIGURE 19-12 Coenocytic bodies in green algae may be more or less isodiametric (spherical) or long, branched, or unbranched tubes. Tubular forms are called siphonous forms.
After sexual reproduction evolved, while all organisms were still unicellular (no multicellular organism is relictually asexual), the simplest life cycle may have been the following: A diploid cell undergoes meiosis, and each cell then exists as a haploid individual, able to undergo mitosis (FIGURE 19-14). Because this is a unicellular organism, cell division is both “growth” and asexual reproduction. Some of these cells can also act as gametes and fuse, producing a diploid zygote that also is able to reproduce asexually—and simultaneously grow—by mitosis. Finally, at some time, some of these undergo meiosis again. Little difference exists between gametes, zygotes, and individuals, none of which is very specialized. Both haploid and diploid cells are capable of growth, division, and reproduction. Such species are dibiontic; that is, there is an alternation of generations between haploid and diploid.

FIGURE 19-13 Individuals of Acetabularia are unusual for being giant single cells, but even more remarkable, they are uninucleate. Each cap is composed of hollow tubes of cytoplasm and vacuole, and at times of reproduction, the sole nucleus begins rapid, repeated mitosis, resulting in hundreds or thousands of nuclei. Cytoplasmic streaming carries them upward into the cap segments; when each segment has several hundred nuclei, it either becomes a dormant cyst and is released or produces hundreds of gametes.
In monobiontic species, specialization occurs in that only one free-living generation exists. In some monobiontic species, the haploid phase represents the individual, and the only diploid cell is the zygote, which is capable only of meiosis, not growth or mitosis (Figure 19-14A and B). The haploid body, either unicellular or multicellular, carries out photosynthesis and growth. In other monobiontic species, the diploid phase represents the individual, vegetative growth phase; the only haploid cells are the gametes, which can undergo only syngamy (Figure 19-14C and D).
In a dibiontic species in which both stages are multicellular, the gametophyte (haploid phase) and sporophyte (diploid phase) may resemble each other strongly, and alternation of isomorphic generations occurs (FIGURE 19-15A), or the two may be very different in appearance and construction (alternation of heteromorphic generations), which allows them to exploit different ecological niches almost as if they were two species; gametophytes do not compete directly with sporophytes (FIGURE 19-15B).
All sporophytes, both in algae and embryophytes, produce spores by meiosis, but many algal sporophytes also produce spores by mitosis; these spores are diploid and grow into a new sporophyte in a form of asexual reproduction. Some algal gametophytes produce spores by mitosis; these are haploid and develop into new gametophytes, also a form of asexual reproduction. In both cases, this is basically similar to asexual reproduction in angiosperms in which some plants produce plantlets along their leaf edges, offset with bulbs or when pieces of cacti break off, root, and act as new plants.
During the earliest stages of the evolution of sexual reproduction, gametes were isogamous (identical), but anisogamy (slight differences in gametes) and oogamy (pronounced either OH-oh-gam-ee or oh-AH-gam-ee) later evolved (FIGURE 19-16). Gametes are produced in gametangia. Sperm cells or microgametes form in microgametangia, and egg cells or megagametes form in megagametangia. Spores are formed in sporangia, either megasporangia or microsporangia, depending on the size of the spore. A critical feature that distinguishes algae from embryophytes is that algal gametangia and sporangia are organized on a cellular level: Individual cells develop into gametes or spores, and when these are released, the gametangium or sporangium is nothing but empty cell walls. In contrast, embryophyte gametangia and sporangia always have one or more outer layers of protective cells, cells that do not turn into gametes or spores; when the reproductive cells are released, one or several layers of complete cells remains.
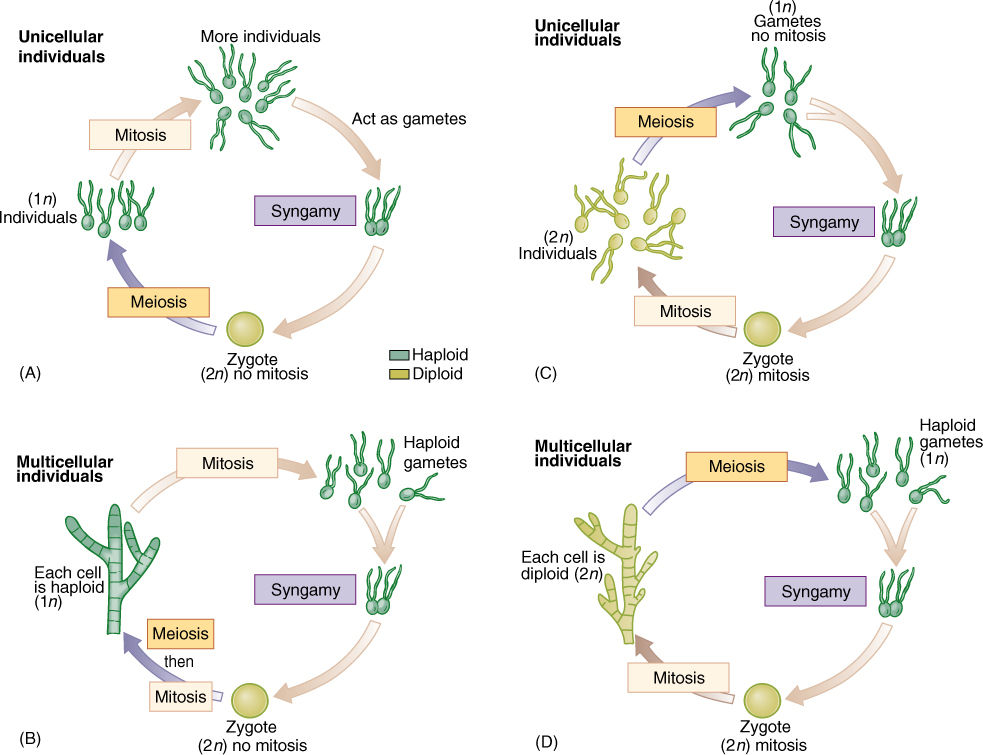
FIGURE 19-14 In monobiontic life cycles, only one generation—one phase—is capable of undergoing mitosis. In most algae (A) and (B), the haploid phase is dominant, undergoing mitosis that results in either more unicellular individuals or a multicellular individual. Only one diploid cell occurs—the zygote—and it cannot undergo mitosis; it cannot produce more diploid cells. (C) and (D) In a few algae and in all animals, the monobiontic life cycle is dominated by the diploid phase, the only haploid cells being the gametes.
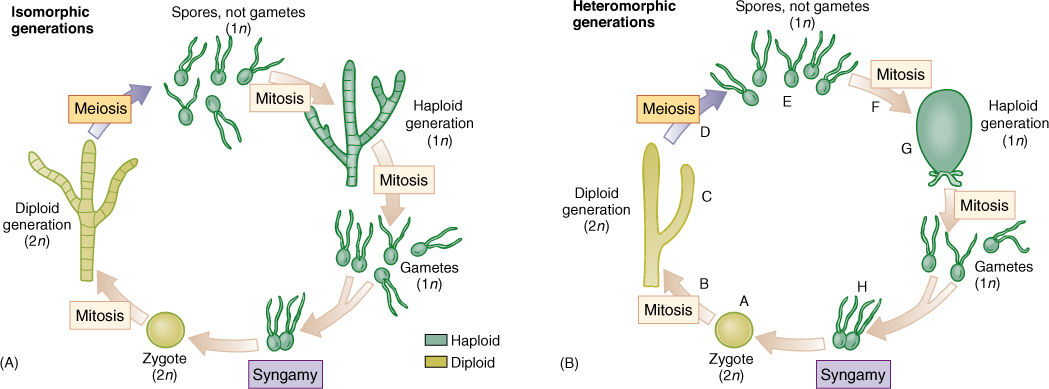
FIGURE 19-15 Dibiontic life cycles involve an alternation of either isomorphic (A) or heteromorphic (B) generations. With isomorphic generations, the similarity of body types can be so great that it is very difficult to know if an individual is a gametophyte or a sporophyte. (B) In dibiontic algae with an alternation of heteromorphic generations, the two types of individuals could never be confused. In flowering plants, the mitosis at B would be the growth of the embryo into the sporophyte plant body, such as an oak tree or a lily, and the meiosis at D would occur only in the sporogenous cells of anthers and ovules. The mitosis at F would be the formation of the tube nucleus, generative nucleus, and sperm cells in pollen and the growth of the megaspore into the antipodals, central cell, synergids, and egg cell of the megagametophyte. The syngamy at H would be fusion of one sperm nucleus with the egg. In flowering plants, the zygote begins growth immediately without a resting period, and flagella are not formed at any stage.
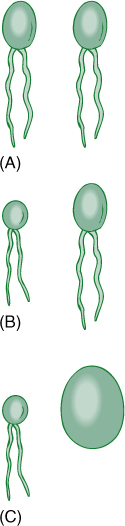
FIGURE 19-16 (A) Isogametes are identical gametes. Theoretically, they could be either motile or nonmotile, but in reality, all known isogametes do have flagella. (B) If gametes have any visible differences but are still similar, they are anisogametes. (C) Oogametes are obviously different in size, and the megagamete is virtually always nonmotile. In most algae and plants, the microgamete has flagella. In flowering plants, conifers, and some algae, microgametes are also nonmotile.
Representative Genera of Green Algae
Unicellular Species
Chlamydomonas is one of the simplest chlorophytes. It is unicellular and, like all green algae, has chlorophyll a and b, carotenoids, and xanthophylls; its starch is formed in chloroplasts just like that of true plants (FIGURE 19-17). It has two anterior flagella that resemble each other; therefore, it is not heterokont like brown algae and their relatives. Like most motile green algae, it has normal mitosis, meiosis, and syngamy. Its life cycle is simple: A haploid cell resorbs its flagella, divides mitotically, and forms 2, 4, 8, or 16 new cells that grow new flagella, each of which swims until it encounters a compatible cell. The cells recognize each other by reactions at the tips of their flagella. They undergo plasmogamy and karyogamy, forming a large zygote that sheds the four flagella and sinks to the bottom in a quiescent state. Germination is by meiosis to form four biflagellate haploid individuals; the zygote is the only diploid cell.
Motile Colonial Species
In the motile colonial line of evolution, cells that greatly resemble Chlamydomonas are produced when the zygote divides, but the progeny cells are held together by a gelatinous matrix (FIGURE 19-18). In Gonium, each colony contains only a few cells (4, 8, 16, or 32), and the only sign of organization is that all flagella beat in a coordinated fashion. Pandorina is about the same size as Gonium but is slightly more derived because it shows a trace of differentiation; the colony swims in one direction, and anterior cells are slightly different from posterior ones. Volvox is the stunning conclusion of this line: Its colonies contain up to 50,000 Chlamydomonas-like cells and are easily visible without a microscope. Differentiation exists in that up to 50 cells in the posterior half of a colony are specialized for reproduction only.
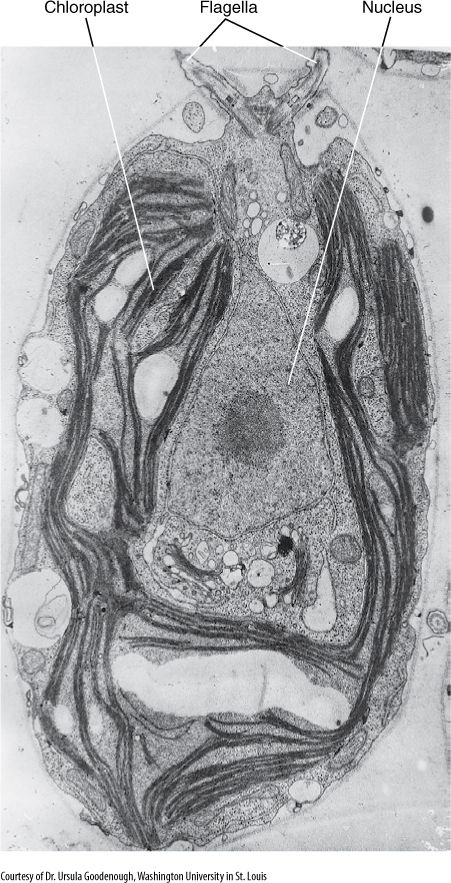
FIGURE 19-17 Chlamydomonas. Its nucleus, chloroplast, and pyrenoid are visible, but only the bases of the two flagella can be seen. The flagella are at the anterior end; that is, they pull the cell forward rather than pushing it. Several starch grains are present.
Filamentous Species
Members of the genus Ulothrix are simple types of filamentous green algae (FIGURE 19-19A). Their life cycle is monobiontic; it involves only one free-living multicellular generation, and it is haploid. It consists of one row of cells that are all rather similar except for the basal cell, which is modified into a holdfast. Filament cells may produce asexual zoospores mitotically; these have four flagella, swim briefly, and then settle and grow into new filaments. During sexual reproduction, some cells produce gametes, which can be identified because they have only two flagella, not four. Ulothrix is isogamous, and the gametes, which all look like cells of Chlamydomonas, pair and fuse. The zygote germinates by meiosis, producing four haploid zoospores, each of which swims for a period, then loses its flagella, attaches to a substrate, and grows into a new filament.
Spirogyra is an extremely common fresh-water filamentous green alga (FIGURE 19-19) found in streams and ponds throughout North America. Its cells have beautiful spiral band-shaped chloroplasts that wind around the cell just below the plasma membrane. Swimming gametes are not formed; instead, filaments undergo conjugation. Each filament is haploid and is either a + or — mating type. If compatible filaments drift against each other, a conjugation tube forms between cells, and the — protoplasts migrate through the tube and fuse with the + protoplasts. Karyogamy follows, although as many as 30 days may pass before nuclei fuse. The cell becomes dormant, thick walled, and resistant; it later germinates and grows into a new filament. Meiosis occurs immediately after karyogamy, and thus, the spore is haploid.
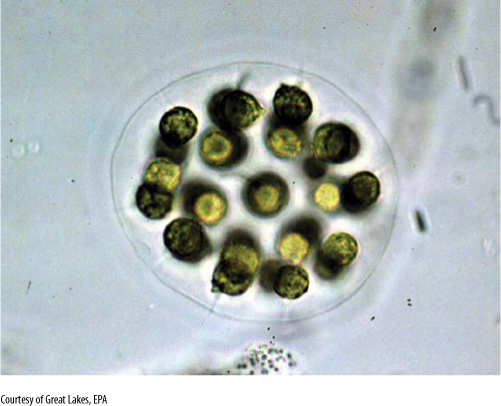
FIGURE 19-18 Eudorina elegans is a small motile colony of green algae. In each species, the number of cells and their spatial arrangements are constant. The numbers and orientations of cell divisions are controlled.
Laminar Species
Ulva is slightly more complex than Ulothrix, but many of its stages are almost identical. A quadriflagellate, haploid zoospore settles down and grows into a Ulothrix-like filament. The cells divide in two directions and thus form a sheet; then all cells divide once in a third direction so that the sheet becomes two layers thick. Ulva has a dibiontic life cycle with an alternation of isomorphic generations (see Figure 19-15A and FIGURE 19-20). During sexual reproduction, gametophyte cells produce biflagellate anisogametes, the smaller gametes being produced on one gametophyte and the larger ones on a different gametophyte. Having two different types of individuals in one generation is dioecy, just as in flowering plants. The zygote grows into a filament and then into a double-layered sheet just like the gametophyte.

FIGURE 19-19 (A) Individuals of Ulothrix are uniseriate, unbranched filaments of haploid cells (×80). (B) Cells of Spirogyra, showing the spiral, band-shaped chloroplasts. (C) Compatible filaments of Spirogyra have been brought together in culture and are undergoing conjugation. This type of plasmogamy is unusual in algae. (D) The yellow material is a green alga, Trentopohlia, growing with lichens and mosses on the surface of a rock in Austin. A rock face in a dry area is not a typical habitat for algae, but Trentopohlia is so well adapted that it not only thrives, it grows large enough to be easily visible. Its green chlorophylls are masked by abundant yellow pigments.
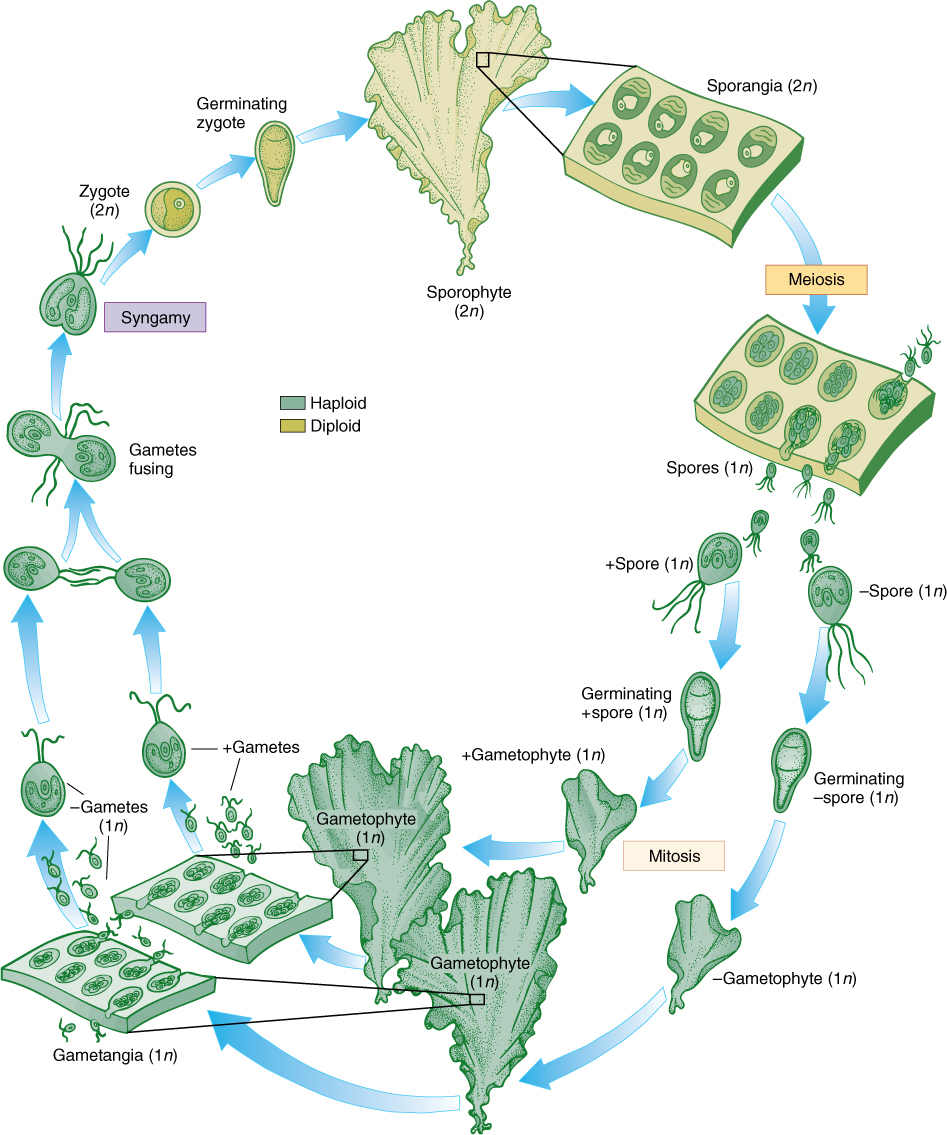
FIGURE 19-20 The life cycle of Ulva. Details are given in text. Two multicellular, isomorphic generations occur. The types of reproductive structures and the size of the nuclei are the only reliable criteria for distinguishing between sporophytes and gametophytes.
Coenocytic Species
A dibiontic life cycle with an alternation of heteromorphic generations is illustrated by Derbesia, the organism in which it was discovered (see Figure 19-15B and FIGURE 19-21). Before going into the details of the life cycle, think for a second about how organisms are studied. Often a biologist goes on a field trip; collects plants, insects, birds, or whatever; and preserves them by drying or fixing them in formaldehyde. It is difficult to maintain things in a living condition. Now imagine trying to do this 100 years ago when transportation was slow, laboratories did not have good artificial light or temperature control, and few pure chemicals were available for making culture solutions. Growing algae to study their life cycles was and is extremely difficult.
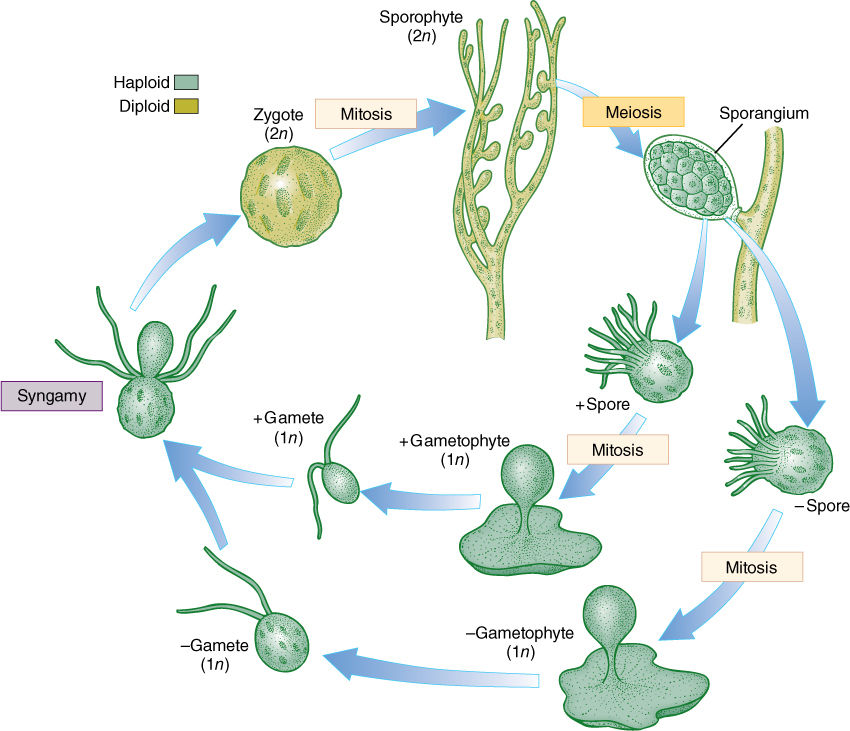
FIGURE 19-21 Derbesia has a dibiontic life cycle with an alternation of heteromorphic generations. The green algae are a large group (approximately 450 genera with 7,500 species), and not all life cycles are known; therefore, many more surprises of this type may be awaiting us. Only by growing all stages of a species can we study its life cycle.
In 1938, P. Kornmann succeeded in obtaining zoospores from Derbesia marina, a branching, filamentous alga made up of giant coenocytic cells (Figure 19-21). The zoospores were carefully maintained, but instead of growing into another Derbesia-like individual as expected, they grew into individuals of a totally different genus, Halicystis ovalis (Figure 19-21). A member of Halicystis looks nothing at all like Derbesia; instead, it is composed of a single large, spherical coenocytic cell attached to the substrate by a small holdfast. Almost its entire volume is one giant vacuole, with only a thin layer of protoplasm next to the wall. When mature, Halicystis individuals, which are the gametophyte stage, produce either male or female anisogametes that undergo syngamy and establish the zygote. After germination, the zygote grows into a Derbesia sporophyte. To classify different life stages as different genera is obviously not correct, and thus, the name Halicystis ovalis has been eliminated. Flowering plants also have an alternation of heteromorphic generations, but the gametophytes grow inside the sporophytes; therefore, the question of which goes with which is easily solved.
Parenchymatous Species
Several groups of green algae undergo a true parenchymatous growth, which is the basis of parenchyma in embryophytes. One group, in chlorophytes, divides by means of a phycoplast, which never occurs in plants, so this group is a remote possibility. The other group, charophytes, undergo cell division by means of a phragmoplast, just as plant cells do. In chlorophytes, the flagellar root apparatus (which anchors flagella to the cell) consists of four bands arranged in a cross; no true plant is known to have this type of root apparatus. In charophytes the flagellar root complex is similar to that of the motile cells of true plants: One major band of microtubules extends down into the cytoplasm from the basal body. Consequently, great attention is being given to charophytes.
An interesting example is Chara (FIGURE 19-22), which has a stem-like body divided into nodes and internodes, with whorls of branches arising at internodes. The body is several cells thick, composed of true parenchyma tissue derived from cell division in all three planes and originating from an apical meristem that contains a prominent apical cell. Although these features seem to correspond to those of flowering plants, virtually all of the resemblance is spurious because the earliest vascular land plants had no nodes, internodes, or branches. If Chara is the sister group to the embryophyte clade, the only features that may be homologous rather than analogous are the parenchymatous body and growth by an apical meristem. Simple parenchymatous bodies also occur in members of Coleochaete, another group of charophytes that are being studied as possible close relatives of land plants.
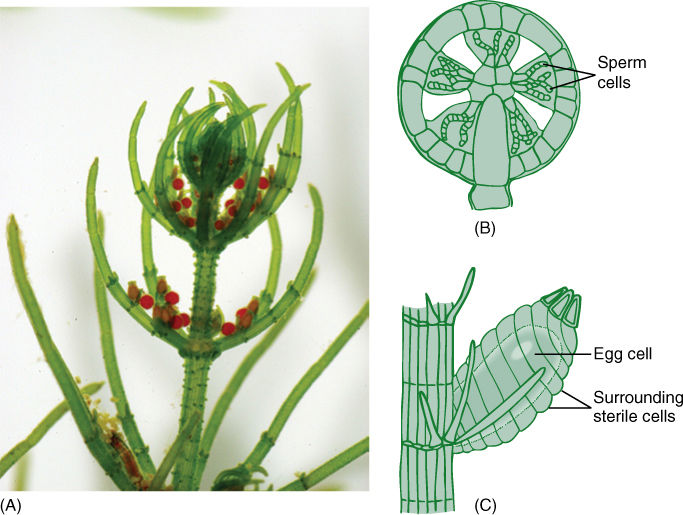
FIGURE 19-22 (A) An individual of Chara, with a parenchymatous body, microgametangia (spherical) and megagametangia (oval). (B) The microgametangium of Chara consists of an outer layer of sterile cells. Some inner cells mature into sperm cells, whereas others remain sterile and act as spacers. (C) The megagametangium is multicellular at maturity but not at initiation. The sterile cells around the egg are not sibling cells of the egg but rather are filaments that grow up from the cell below the egg. This is a multicellular gametangium, but it is filamentous, not parenchymatous.
Reproduction in Chara is significant. Embryophytes have multicellular reproductive structures with sterile cells, and on this basis, Chara would have to be classified as a plant rather than an alga. Its sperm cells are produced in a truly multicellular gametangium whose outer cells are sterile; only the inner cells convert to sperm cells (Figure 19-22B). At maturity, the outer cells separate slightly and the motile sperm cells swim away. The egg is formed as the terminal cell of a short filament three cells long, but the subterminal cell subdivides, and those cells grow upward and surround the egg (Figure 19-22C). After fertilization, the sterile cells surrounding the fertilized egg deposit thickenings on their inner walls, those adjacent to the zygote. The resting structure thus consists not only of a thick-walled zygote but also protective sterile cells. When the resting cell germinates, it grows out as haploid filaments that soon establish an apical meristem and parenchymatic growth.
Green Algae and Embryophytes
Figure 19-3 shows charophytes and embryophytes as sister groups sharing a common ancestor. As such, they form a monophyletic clade, and some people have suggested that this clade be named the streptophytes, or even that the definition of embryophytes should be extended, that charophytes should be considered true plants, not algae. Most people hesitate to establish new formal classifications for clades and nodes; consequently, the charophyte + embryophyte clade is referred to with the informal name streptophyte.
Notice that the phylogenetic hypothesis depicted in Figure 19-3 also proposes a monophyletic clade that consists of red algae + green algae + embryophytes, and glaucophytes could be added as well. Names have been suggested for this group also, the archaeplastids (because this clade is based on the original plastid endosymbiosis) or primoplantae. So far, these names have not been used very often.
![]() Red Algae
Red Algae
The red algae constitute a large group (approximately 400 genera and 3,900 species) of especially distinct and fascinating algae (see Figure 19-2H, FIGURE 19-23, and FIGURE 19-24). Numerous structural, biochemical, and reproductive features set them off from other algae as well as from embryophytes. One of the most important biochemical distinctions is that like cyanobacteria they contain phycobilin accessory pigments that are aggregated into phycobilisomes (see Figure 19-6). Their red color is due to the presence of phycoerythrin; however, they are often purple, brown, or black because of the additional presence of phycocyanin, just as in cyanobacteria. Carotenoid accessory pigments are also present, as is chlorophyll a. The actual quantities of each type of pigment vary with depth. Those algae that grow in bright surface waters have an array of pigments suitable for relatively intense light, whereas algae in deeper waters have a different complex of pigments better suited both to dim light and to the altered spectrum present as a result of the water’s differential color absorption.
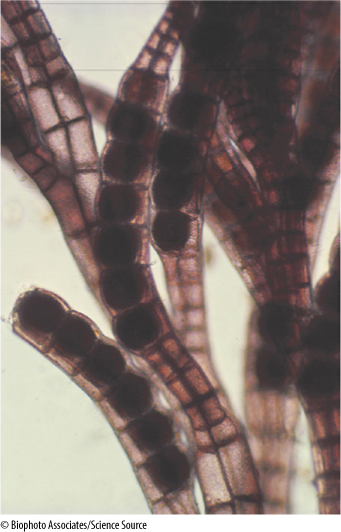
FIGURE 19-23 Many red algae have highly branched, filamentous bodies. Polysiphonia (×20).

FIGURE 19-24 Coralline red algae have walls so heavily impregnated with calcium carbonate that the body is hard and brittle.
Excess photosynthate is stored as floridean starch, a branched polymer of glucose somewhat similar to glycogen; it occurs as granules in the cytoplasm, never in the chloroplast. Other reserves occur that contain unusual sugars such as floridoside and isofloridoside, indicating that the carbohydrate metabolism of red algae has characteristic features not shared by plants.
In addition to a thin layer of cellulose, walls of red algae contain a thick layer of slimy mucilages called sulfated galactans. These are important commercially as thickening, suspending, or stabilizing agents in puddings, ice creams, cheeses, and salad dressings. The culture medium agar is also extracted from them. The complete wall—cellulose layer plus mucilage—is quite thick, often as thick as the protoplast is wide. Almost all red algae are multicellular, and thus, a large fraction of the individual’s volume consists of this apoplastic space. As a result, all cells, even the most internal ones, may have relatively direct contact with the surrounding water. In the largest family of red algae, the Corallinaceae or coralline red algae, such large amounts of calcium carbonate are deposited into the walls that they become rocklike (Figure 19-24). It was not until 1837 that they were recognized as algae rather than corals.
Walls of red algae lack plasmodesmata of the type that occurs in plants, but they do have distinctive pit connections. During cell division, the new wall is formed as a ring that grows inward from the original cell wall; the growth of the septum stops before it is complete, leaving a hole in the center. Vesicles then deposit a material that precipitates, forming a lens-shaped plug. Whether these are a means of intercellular transport or communication is not known, but when two separate filaments of cells come into contact, a “secondary pit connection” can form between them.
Red algae are usually multicellular; only a few unicellular species (Porphyridium, Rhodospora) have ever been discovered. Most red algae tend to be rather conspicuous, often beautiful individuals, and are filamentous, membranous, or foliaceous; true parenchymatous bodies are reported to occur in only a few algae, such as Bangia and Porphyra. Typically, red algae are smaller and less complex than brown algae.
Despite the rather large size that bodies of red algae can attain, little differentiation or specialization occurs among the cells. The greatest differentiation typically involves only cell size and pigmentation. Outer cells are smaller and more heavily pigmented; inner cells are larger and have fewer, smaller chloroplasts. Most species grow attached by rhizoids to rocks, shells, algae, or sea grasses. Numerous red algae (more than 40 genera) are parasitic, usually on other red algae. The basal cells of these species penetrate into the host, forming secondary pit connections with host cells.
Life cycles of most red algae are poorly known, but the few that have been studied well are all extremely complex, almost all involving at least one multicellular stage but none having any motile cells. Flagella and centrioles do not occur in any stage of any species. Many variations occur, and there is no “typical” red alga life cycle; FIGURE 19-25 is a generalized life cycle. Gametophytes bear gametangia that produce nonmotile sperm cells called spermatia and egg-like cells called carpogonia. Carpogonia are large cells with a long tubular extension that basically acts as a receptor for drifting spermatia. When a spermatium contacts the extension, plasmogamy occurs and the nucleus migrates to the carpogonium base, where karyogamy occurs. In any other group, this cell would be a zygote and would either grow or produce spores, but in many red algae, the fertilized carpogonium puts out another long filament that carries the diploid nucleus out of the carpogonium and deposits it into a totally different cell, an auxiliary cell, where mitosis begins and produces a mass of cells. This mass is a new generation, the carposporophyte, and it produces diploid carpospores by mitosis. The carposporophyte is a totally distinct generation that has no equivalent in any other algae or plants. Carpospores are released, float away and settle, and then germinate and grow into diploid individuals called tetrasporophytes, equivalent to a regular sporophyte. These have sporangia in which the cells undergo meiosis, producing haploid tetraspores that grow into gametophytes.
![]() Brown Algae and Their Relatives: The Heterokonts
Brown Algae and Their Relatives: The Heterokonts
Brown Algae
Brown algae are almost exclusively marine; only a few fresh-water species are known (see Figure 19-2G and FIGURE 19-26). They prefer cold water that is very agitated and aerated. They can be found most easily on rocky coasts growing in the littoral zone, the region between low tide and high tide, also called the intertidal zone, where they are periodically exposed to air and full sunlight. Over 1,500 species are known, grouped into about 250 genera. Anatomically and morphologically, brown algae have the most complex bodies of any algae; some are considerably more complex than plants such as mosses and liverworts. Although very distinct from embryophytes biochemically and ecologically, the two groups have remarkable parallels in the types of bodies and life cycles that have evolved.
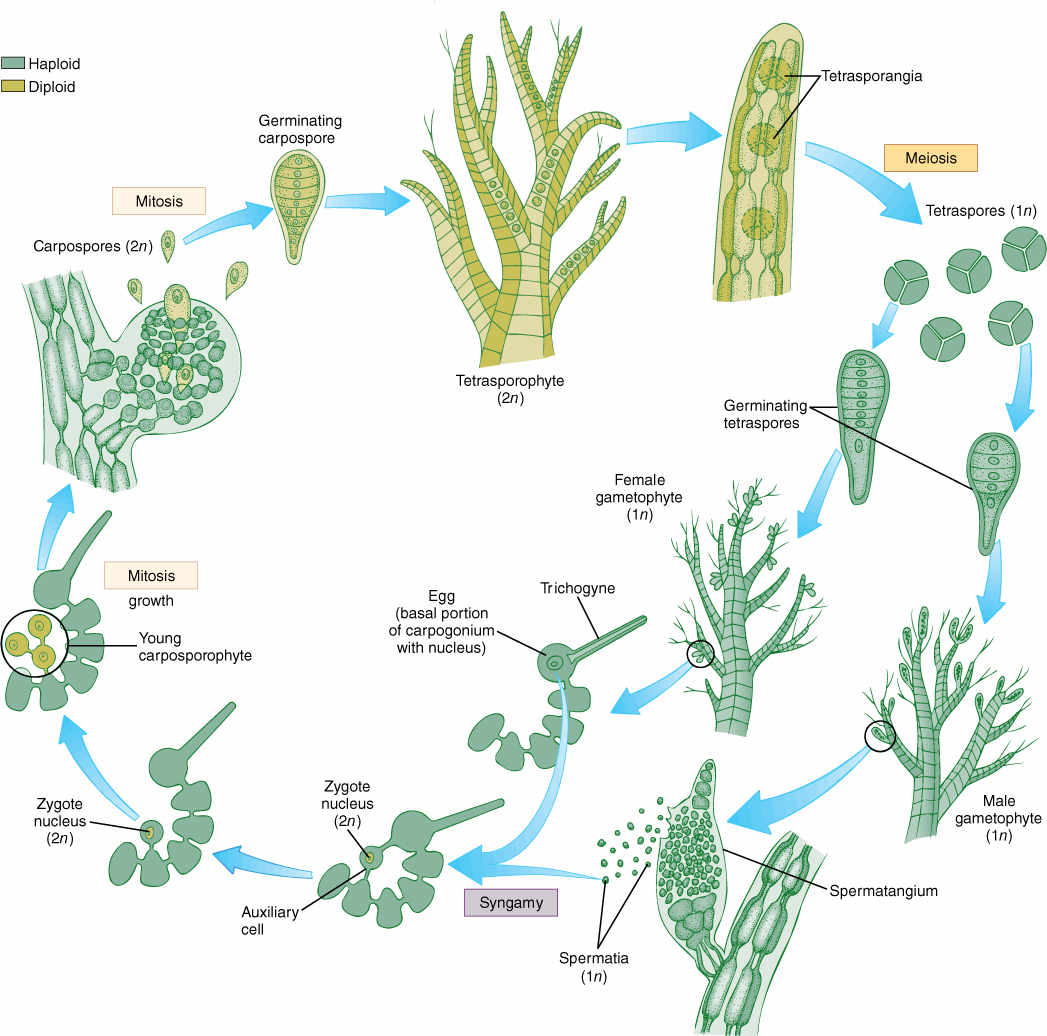
FIGURE 19-25 The life cycle of a red alga. Details are given in text. Note especially the lack of swimming cells and the presence of three generations, one haploid and two diploid.
The differences between brown algae and green organisms (both green algae and plants) are clear cut. Brown algae have chlorophyll a and chlorophyll c and large amounts of a variety of pigments such as fucoxanthin, violaxanthin, and diatoxanthin (these are xanthophyll pigments). Carotenes are also present. Their suite of pigments permits brown algae to carry out photosynthesis at numerous levels in the ocean. Sunlight differs not only in intensity but also in quality at different depths: White light with a full spectrum occurs at the surface, but primarily just blue-green light reaches depths of 50 meters or more. Membranes of brown algal chloroplasts associate into grana-like stacks, but they are always small, each consisting of just two or three membranes (see Figure 19-8).
The storage product of brown algae is laminarin (a polymer of glucose), mannitol, or fats, but not starch. Laminarin constitutes up to 34% of the body weight of some brown algae. Most other algae, especially the marine species, live in such a stable environment with regard to light, temperature, and nutrients that photosynthesis and growth occur more or less continuously, so large reserves are unnecessary; however, many brown algae live in seasonal habitats, and they need energy reserves. For example, kelps have large bodies made up of three parts: holdfasts and stipes (stalks) that may be perennial (some live at least 17 years) and photosynthetic blades that are annual (FIGURE 19-27). When blades become moribund and decompose, the holdfasts and stalks must subsist on stored nutrients until the new blade can be formed and begin photosynthesis in the spring.

FIGURE 19-26 Fucus is a common brown alga you might see on any cool, rocky coast, just below the high tide level. It tolerates being exposed to air for several hours at low tide because its thick cell walls are extremely hydrophilic, slowing the evaporation of water.
Cell walls of brown algae contain cellulose and alginic acid, an unusual polymer of D-mannuronic acid and L-guluronic acid not found in other algae. The alginic acid component of the wall is gummy or slimy and causes the filaments of cells to adhere into a compact body. It may have another unknown function, however, because as much as 24% of the body dry weight can be alginic acid in Ascophyllum.
A remarkable feature of brown algae is that they are all multicellular; no unicellular species is known to exist, and individuals of many species of kelps become huge and complex. One of the simplest brown algae is Ectocarpus. It has an alternation of isomorphic generations, both generations consisting of uniseriate branched filaments that arise from a prostrate branched system attached to rocks, shells, or other, larger algae. In the diploid sporophytes, some of the terminal cells of small lateral branches enlarge greatly and become sporangia. The nuclei divide repeatedly, and then 32 or 64 zoospores are released. The first nuclear division is meiotic, and the spores are therefore haploid. After swimming temporarily, zoospores settle down and grow into gametophytes that are almost identical to the sporophytes. The primary difference is that at the ends of the branches are multicellular gametangia, not unicellular sporangia as in sporophytes. The gametes are anisogamous: Some settle and attract others by secreting a sex hormone ectocarpene. After fertilization, the zygote grows into a new sporophyte.
A more complex brown alga is Fucus, which is common on rocks in the intertidal zone (see Figure 19-26 and FIGURE 19-28). The diploid individuals are exposed at low tide, and their large dichotomously branched bodies (up to 2 m) are attached to rocks by holdfasts. The bodies are complex histologically with epidermis, cortex, and a central region. The ends of the branches are called receptacles and are swollen with large deposits of hydrophilic compounds. Scattered over the surface of the receptacles are minute openings that lead to small cavities, conceptacles; some conceptacle cells undergo meiosis, producing either large eggs or small sperms. At low tide, individuals are exposed to air, and the conceptacles contract, squeezing out gametes. When the tide comes in, gametes are washed free and fertilization occurs in the water. Fertilized eggs settle to the bottom and grow into new diploid individuals. No free-living haploid generation occurs; Fucus is monobiontic.
Individuals of the kelp Nereocystis that measure up to 45 m—taller than most trees—are not uncommon, but they are not bulky, being less than 5 cm thick. Any alga that becomes as long as half a football field must have specialized regions to its body. The holdfasts of a kelp are located in deep, dark, poorly aerated water, whereas their blades exist in brightly lighted and well-aerated surface waters. To keep from being damaged by wave actions, the bodies must be firm, elastic, and thick—too thick for diffusion alone to mediate exchange of gases, nutrients, and wastes. Some species have an epidermis-like outer covering, called a meristoderm, which adds new layers to the surface. Interior to the meristoderm is a parenchymatous middle tissue that resembles plant cortex, and in the center is a cylinder of trumpet cells that resemble sieve tube members (FIGURE 19-29). Trumpet cells carry out long-distance transport of carbohydrates through the body. They are elongate, have large holes that occur in a sieve-like arrangement in the end walls, and the holes are lined with callose. If radioactive carbon dioxide is given to the blade, a short time later radioactive photosynthates can be detected in trumpet cells of the stipe. The flow rate can be as high as 65 to 78 cm/hr. Like sieve tube members, trumpet cells function only temporarily and then are replaced by new ones that differentiate from cortex cells.
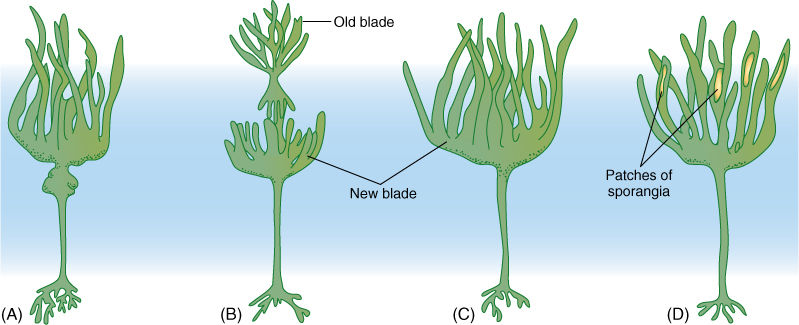
FIGURE 19-27 (A)-(D) In many kelps, the blade is annual but the stipe and holdfast are perennial. A new blade is formed each year; if the old blade has not been completely destroyed by wave action, it is sloughed off.
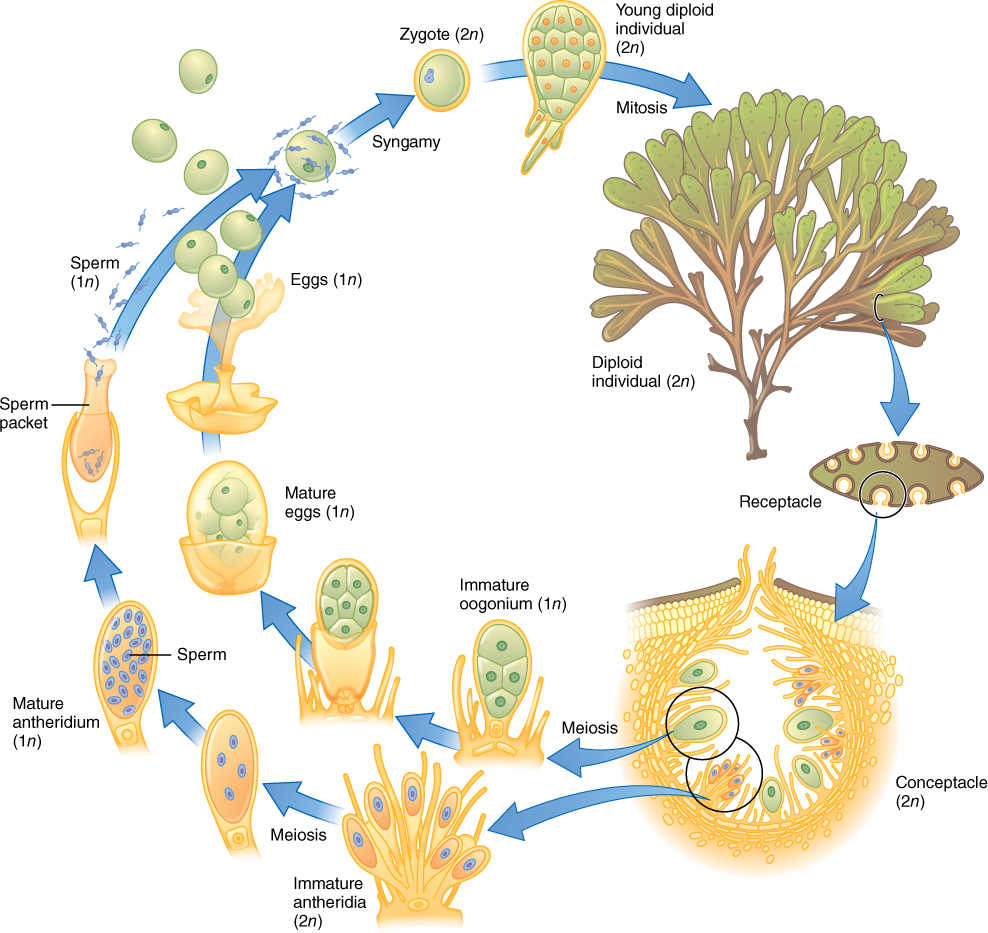
FIGURE 19-28 The life cycle of Fucus. Details are given in text.
Although trumpet cells and sieve tube members are extremely similar structurally and metabolically, they are not at all homologous. The most recent common ancestor shared by brown algae and angiosperms must have been a unicellular nonphotosynthetic protist that could not possibly have had either trumpet cells or phloem. This is an outstanding example of convergent evolution (a plesiomorphy) of tissues.
The junction between an air bladder (pneumatocyst) and a blade is an intercalary meristem capable of prolonged growth, which produces blades several meters long. Growth can be extremely rapid, up to 6 cm/day. Blades are photosynthetic and vegetative for a period after their formation by the intercalary meristem, but then portions become fertile and produce haploid zoospores by meiosis. Under inductive conditions, sporangia form either in patches or over the blade’s entire surface. In Macrocystis, some blades remain vegetative and others are specialized and produce sporangia. Macrocystis releases as many as 76,000 spores/min/cm2 of reproductive blade surface; the mean rate over long periods is 5,000 spores/min/cm2. Spores grow into tiny, filamentous gametophytes that somewhat resemble small plants of Ectocarpus—an alternation of heteromorphic generations. Gametophytes produce oogametes, and after fertilization, a new sporophyte is formed.
FIGURE 19-29 (A) A longitudinal section through the stipe of the kelp Macrocystis. In the center are numerous trumpet cells (×290). (B) A young trumpet cell, showing the enlarged end walls where holes have developed (×500).
Diatoms
This clade of heterokont algae contains about 200 genera and 5,000 species. Diatoms are easy to recognize because of their distinctive morphology. Each cell has a wall composed of two halves or frustules that fit together like a Petri dish and its lid (see Figure 19-2C and FIGURE 19-30). Each frustule is encrusted with silica, and when the diatom dies and the protoplasm degenerates, the siliceous frustules sink to the ocean floor and accumulate. Such deposits, known as diatomaceous earth, can become hundreds of meters thick and cover many square kilometers. Accumulations of this volume are possible because diatoms are the most abundant organisms in the oceans, and a single liter of seawater may easily contain more than 1 million individuals.
It might be thought that organisms consisting of a single cell could not possibly be very complex and certainly would not have enough structure to allow 5,000 species to be recognized, but frustules are extremely intricate. Where the two come together, they are held in place by girdle bands, and each species has its own characteristic complicated pattern of ridges, depressions, and pores. The cells are either round in face view (centric diatoms) or elongate (pennate diatoms).
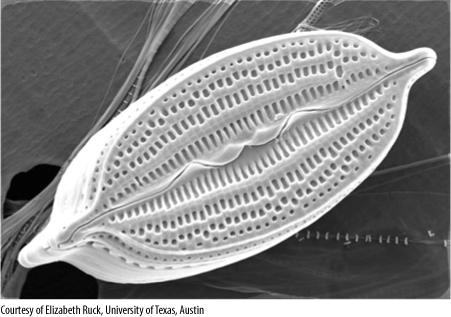
FIGURE 19-30 Pennate diatoms are elongated and bilaterally symmetrical. The numerous holes in their frustules allow nutrients to be taken up from seawater and wastes to be excreted (×4,500).
When a diatom undergoes mitosis and cytokinesis, each progeny cell receives one of the frustules from the parental cell, using it as its outer frustule and synthesizing a new inner frustule. All cells that receive the inner frustule automatically mature into a cell that is smaller than the parent cell, and thus, the average cell size decreases with each cell cycle. This obviously cannot go on forever, and when a cell reaches a critical small size, sexual reproduction is triggered. The cells undergo meiosis, some producing 4, 8, or 16 sperm cells, others producing just one or two large egg cells. After fertilization, the zygote grows into a large cell and then becomes dormant, surrounded by a wall different from that of a vegetative cell. The zygote finally “germinates,” undergoing a round of nuclear and cell division, and reinitiating the process of decreasing cell size.
Yellow-Green Algae
Yellow-green algae occur mostly in fresh water, and formerly, many were thought to be green algae until chlorophyll c was discovered in them (see Figure 19-2E). They are somewhat diverse, some being unicellular, some filamentous, and some forming giant multinucleate cells. Filamentous forms, such as Tribonema, have walls like diatom frustules, each cell having two half-walls, but each of these frustule-like walls is firmly attached to the adjacent half-wall of the neighboring cell. Although daughter cells remain together after cell division, the organism can be called multicellular only in the broadest sense because no differentiation or specialization occurs in any part: The filament is really just a colony of individuals that happen to remain together.
Some yellow-green algae, such as Vaucheria, have an unusual body that consists of a long tubular coenocyte with a single large central vacuole, a thin peripheral layer of cytoplasm, and thousands of nuclei. During asexual reproduction, the cytoplasm at the tip of a tube is isolated by a transverse wall, and pairs of flagella form near each nucleus; then the old wall breaks open and the multiflagellate “cell” swims slowly away (FIGURE 19-31). It soon loses its flagella and begins growth at each end, forming a new tube. The greatest complexity is displayed during sexual reproduction. A mass of cytoplasm is walled off and pairs of flagella form, but then each nucleus organizes the protoplasm around it into an individual cell. When the original wall breaks down, numerous biflagellate sperm cells swim away. An egg cell is formed as a small multinucleate protrusion on a tube, but as the cross wall forms, all nuclei except one migrate out, leaving a single large uninucleate cell. After fertilization the zygote forms a thick wall and is abscised from the parent branch. After a period of dormancy, the zygote undergoes meiosis and germinates, forming a new tube filled with haploid nuclei.
Golden-Brown Algae
Golden-brown algae consist of about 70 genera and 325 species. They are single cells covered with numerous tiny siliceous scales that develop within special vesicles in the endoplasmic reticulum (see Figure 19-2D and FIGURE 19-32). Cells may be either uniflagellate or biflagellate; rarely they have no flagella, crawling by amoeboid motion instead. Although they are photoautotrophic because of their chloroplasts, they can also ingest bacteria by phagocytosis.
Golden-brown algae are particularly important because of their past biology. The living cells float in warm, sunny surface waters, like diatoms, but when they die, the lack of swimming motion allows their dense siliceous scales or frustules to sink and accumulate on the ocean floor as chalk (blackboard chalk used to be made from this). The remains of both diatoms and golden-brown algae, especially a group known as coccolithophorids (usually called coccoliths), are relatively inert and do not decompose. Drilling into sea floor sediments makes it possible to analyze the amount of diatom and coccolith remains and to estimate the rate at which they accumulated. It is possible to reconstruct the surface climate. Accumulation was rapid when waters were warm and carbon dioxide was abundant for photosynthesis and slow when the climate was cool or if carbon dioxide was low. Shallow coastal seas tend to be warmer than the open oceans, and this also affects coccolith growth and reproduction. By also analyzing fossil animals, which are affected by temperature but not carbon dioxide, we can reconstruct past climates and ocean basin geology precisely.
![]() Dinoflagellates
Dinoflagellates
Dinoflagellates have many unusual characters (see Figure 19-2B and FIGURE 19-33). Their nuclear envelope and nucleolus persist throughout mitosis, and a typical spindle does not form. Instead, channels open in the nucleus and large bundles of microtubules pass through. Their chromosomes, which do not have centromeres and are permanently condensed, are attached to the nuclear envelope, whose expansion during the formation of the nuclear channels causes the chromosomes to split. There are no histones.
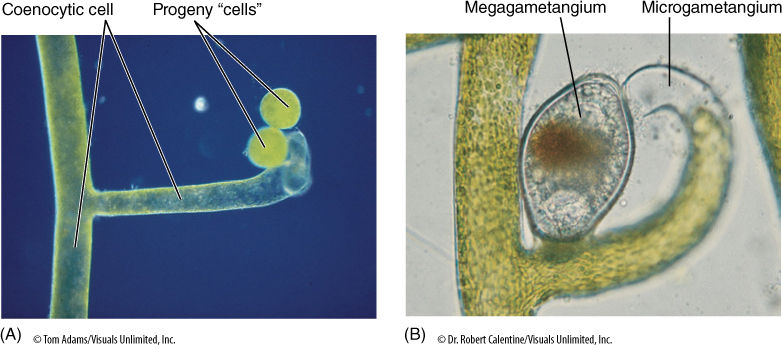
FIGURE 19-31 (A) Vaucheria occasionally walls off the tip of a coenocyte; then pairs of flagella form near each nucleus. The wall breaks open, and the “cell” swims away, settles, and grows into a new individual (×100). (B) During sexual reproduction, one segment of the coenocyte forms numerous biflagellate sperm cells, and another forms one large uninucleate egg. After the sperm cells swim away, all that is left of the gametangium is wall; unlike gametangia in true plants, no protective layer of sterile cells is present (×200).
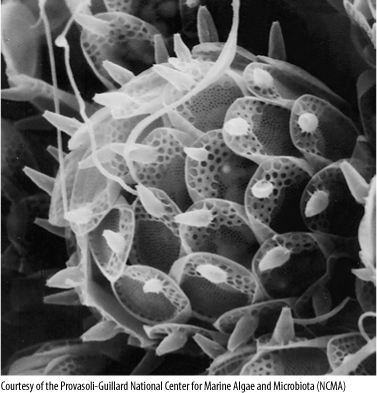
FIGURE 19-32 Each cell of a golden-brown alga is covered with many minute scales that are formed within vesicles of the endoplasmic reticulum. The pattern within each scale is constant for each species, but how it is controlled is unknown. Synura uvella (×5,000).
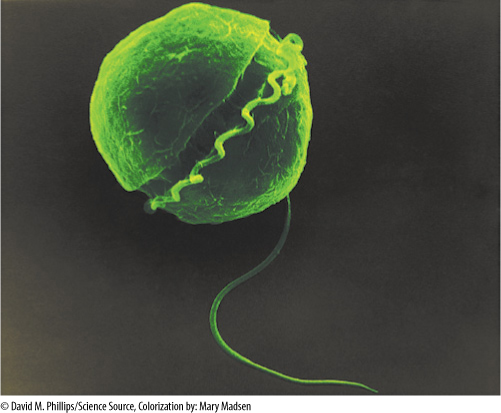
FIGURE 19-33 Dinoflagellates have two flagella, one of which lies in a groove that encircles the cell (×10,000).

FIGURE 19-34 The population density of dinoflagellates can rise so high that they color the water, producing a red tide. A few centimeters away, the density is so low that the water appears normal.
Dinoflagellates are almost exclusively motile and unicellular; only a few species such as Gonyaulax monilata form chains of similar cells with no differentiation. Their flagella have a strikingly characteristic arrangement in dinoflagellates: One long flagellum lies in a longitudinal groove with its distal end free and responsible for swimming. The other flagellum is flat and ribbon-like and lies completely within a transverse groove that encircles the cell. They differ from heterokonts by not having a tinsel flagellum. Most species are photosynthetic, but many are completely heterotrophic. Dinoflagellates have chlorophylls a and c, not a and b, and their carotenoids are quite unusual. In addition to β carotene, they also have several unique xanthophylls such as peridinin and dinoxanthin. Their reserve material is starch or oil. Dinoflagellates may lack a wall or have one consisting of cellulose plates whose arrangement and number are useful for identifying genera and species.
During dinoflagellate sexual reproduction, vegetative cells release small, naked cells that act as gametes. At the initiation of fusion, they become quiescent but then resume swimming, and the zygote may remain motile for 12 to 13 days. It then becomes thick walled and quiescent for approximately 2 months. During germination, it undergoes meiosis and a single haploid vegetative cell emerges—a new individual that grows and multiplies by mitosis.
Organisms with many relictual characters are often regarded as poorly adapted, perhaps surviving only in certain specialized, protected environments, but dinoflagellates are abundant throughout the oceans. Under conditions of ideal temperature and nutrients, population growth of some dinoflagellates, especially Gonyaulax and Gymnodinium, is explosive. Within a few days they become so numerous that their bodies actually color the water reddish brown—a “red tide” (FIGURE 19-34). The density of dinoflagellates can be as high as 30,000 cells per milliliter of sea water. Red tides are becoming increasingly frequent in the Gulf of Mexico, a phenomenon that is unexplained at present. This fascinating biological spectacle is also dangerous because these algae produce toxins, and their large concentrations kill fish and make other marine life poisonous to humans. The poisons of many species, Gonyaulax catenella, for instance, are potent neurotoxins, interfering with the movement of sodium ions across our nerve membranes.
Oomycetes
These protistans were thought to be fungi for many years. They have no chloroplasts, and thus, like fungi, they are never photosynthetic. Also, many of them have bodies that consist of long, slender multinucleate, coenocytic tubes that have no cross walls, an organization typical of fungi; however, many of their characters are more like those of algae and plants but different from true fungi. For example, oomycetes have a cellulose wall but all true fungus cell walls are based on a polymer called chitin (as occurs in animals). Also, oomycetes have tinsel flagella, and their chromosomes are fairly large (chromosomes in fungi are extremely tiny). DNA sequences now strongly support inclusion of these groups in the heterokont protists.
Oomycetes are diverse in structure and nutrition. Lacking chloroplasts, they must be parasitic or saprophytic. Most, such as Saprolegnia and Achlya, are aquatic and have been called “water molds,” but others live in dry habitats such as on and within leaves. Some consist of single cells that live as parasites inside rotifers, minute aquatic animals. Many oomycetes, however, form extensive networks of slender tubular cells.
Sexual reproduction occurs in most oomycetes: A cell swells into a bulbous oogonium, which contains one or several diploid nuclei that undergo meiosis, each in its own mass of protoplasm (FIGURE 19-35). An adjacent cell elongates as an antheridium and contacts the oogonium; a hole forms, and microgametes pass directly into the oogonium, fertilizing the megagametes. The resulting oospores are diploid and are actually zygotes; each matures into a resting spore with a thick cellulose wall. These remain viable for many months even under dry, adverse conditions. Under favorable conditions, they germinate into diploid tubular cells.
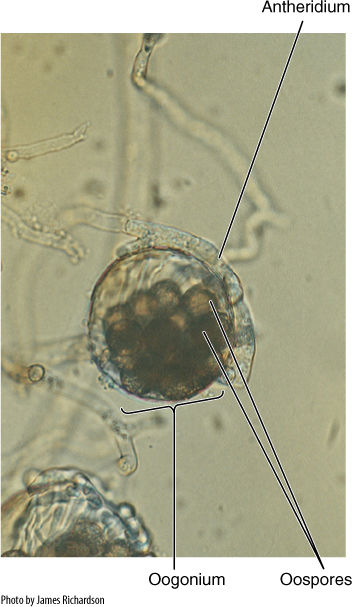
FIGURE 19-35 Saprolegnia: An antheridium has already contacted the oogonium and transferred microgametes. Oospores have formed. This situation is a form of oogamy (×3,500).
Two oomycetes (Plasmopara viticola and Phytophthora infestans) are severe plant pathogens. The first attacks grapes and was introduced to France in the late 1870s on grafting stock imported from the United States. Plasmopara viticola caused tremendous damage throughout vineyards, except along roadsides where a mixture of lime and copper sulfate had been applied to the plants to keep passersby from stealing grapes. After it was recognized that this inhibits the pathogen without damaging the grape plants, it was used for treating entire vineyards.
Phytophthora infestans, an oomycete disease of potatoes, caused the great potato blight and famine in Ireland in 1846—1847. At that time, Ireland was overpopulated, and the potato was almost the only food for most people. The infection with P. infestans spread so rapidly and virulently that virtually the entire food crop for the country was destroyed; as many as 800,000 people starved to death the following winter, and more than 1 million emigrated, mostly to the United States. Phytophthora infestans is still a serious disease, and in cool, moist climates, it is controlled only by regular spraying with algicides.
Hypochytridiomycetes are a related group of nonphotosynthetic protistans that were formerly thought to be fungi.
![]() Euglenoids
Euglenoids
More than 800 species of euglenoids have been discovered, named, and placed into 36 genera (Figure 19-2A and FIGURE 19-36). The majority (25 genera) never have chloroplasts at all, but 11 do, derived through a secondary endosymbiosis with a green alga. Most euglenoids are unicellular, but in some species, a few cells remain together after cell division.

FIGURE 19-36 Euglena: The pellicle, now often called a periplast, is not a cell wall, but rather a thick layer of elastic proteins that accumulate just below the plasma membrane (×11,700).
Green, photosynthetic euglenoids have chlorophylls a and b (just as do green algae), but carbohydrate is stored as the glucose polymer paramylon, not as starch. Euglenoid chloroplasts are extremely sensitive to antibiotics that interfere with 70S ribosomes, and exposure to streptomycin results in the loss of chloroplasts; colorless “bleached” individuals can be produced with this treatment. Many colorless individuals, both natural and bleached, take in food particles by phagocytosis (engulfing their food), and most absorb sugars and even proteins through their plasma membrane. Even chlorophyllous species grow faster in light if supplied with organic compounds, and all must obtain B vitamins from the environment.
Euglenoids swim actively with two flagella located at the cell’s anterior end, one short and the other quite long. Euglenoids differ from other protists and plants in an important character: They never have a cellulose wall. Instead their surface contains a layer of elastic proteins that constitute a pellicle (also called a periplast), which is located under the plasma membrane and gives the cell some rigidity and shape.
Euglena is a representative example. Individuals are elongate cells that, because of their pellicle, have a somewhat ovoid shape, although they flex as they swim. At the anterior end is an invagination, the canal. The two flagella are attached at the base of the canal and extend upward, but only one is long enough to emerge from the canal and be useful for swimming. At the side of the canal, near the base of the flagella, is an eyespot composed of many small, orange droplets of carotenoids. There are numerous disk-shaped chloroplasts, each containing a region called a pyrenoid; in most algae, the pyrenoid is involved in polymerizing sugars into reserve polymers. That may also be true in Euglena, but the reserve paramylon accumulates outside the chloroplast, away from the pyrenoid.
Euglenoids have many features that must have undergone little change and thus probably are similar to those present in early eukaryotes. During mitosis, their nuclear envelope and nucleolus persist, and spindle microtubules form within the nucleus. Chromosomes are aligned lengthwise on the spindle, not crosswise as in other eukaryotes, and each chromatid appears to have its own minispindle consisting of four centromere microtubules and 8 to 12 longer microtubules. Cell division is by longitudinal cleavage, not by either a phragmoplast or a phycoplast, and after division is completed, chromosomes remain condensed. Neither meiosis nor sexual reproduction has been found in any euglenoid. Many features of normal mitosis and meiosis as occur in plants and animals apparently did not arise until after the euglenoid clade had become reproductively isolated from the rest of the early eukaryotes.
 At the Next Level
At the Next Level
1. Complex multicellular bodies in algae. Both red algae and brown algae have complex bodies composed of several distinct tissues. Each evolved separately (separately from each other and from true plants), so each presents many new alternatives for constructing a complex sessile body (a body attached to a substrate, one that cannot move away). Both have complex meristematic and reproductive tissues, and many brown algae conduct sugars through tissues that resemble phloem but which originated independently: It is the result of convergent evolution with phloem. Studying these complex algal bodies will give you a more complete appreciation for the bodies of true plants.
2. Red tides. In many cases, organisms become more complex as they evolve to become more adapted. But this is not always true. Many of the unicellular algae are so well-adapted to ocean habitats that nothing outcompetes them, and when conditions are just right, unicellular algae become so abundant they cause red tides. Some of the red tide dinoflagellates are poisonous and kill animals in the area, altering the community biology. As we put more sewage and fertilizers into rivers that empty into oceans, and as global temperatures rise, red tides are becoming more frequent and may have a permanent effect on coastal ecology.
3. Acetabularia: complex giant cells with a single nucleus. Many algae have unicellular bodies, but species of Acetabularia are spectacular. Each cell is gigantic, almost 1 cm long, and often have a stalk, a cap, or finger-like extensions, and despite the complexity of such cells, they have just one nucleus. Many ingenious experiments were performed in the early days of developmental biology (the mid-1900s) in which nuclei were transplanted from a cell with one shape into the enucleated body of a cell that had a different shape. After a short recovery, the cells would change their shape to correspond to the instructions from the new nucleus. At present, we take such things for granted, but at that time this was extraordinary proof of the importance of nuclei. Reading about the research performed with Acetabularia will expand your appreciation of the ingenuity and talent needed to discover the facts we rely on today.
SUMMARY
1. Several significant evolutionary divergences occurred in the early eukaryotes, long before the plant, animal, and fungus clades had originated. Euglenoids have no meiosis or sexual reproduction. Dinoflagellates lack histones and normal mitosis; several groups have surfaces that lack cellulose or have cellulose combined with chemicals not found in plants.
2. A primary endosymbiosis produced the chloroplasts of glaucophytes, red algae, green algae, and plants. Several events of secondary endosymbiosis of entire red or green algae produced chloroplasts of the other protists.
3. Algae are distinguished from plants (embryophytes) by having gametangia and sporangia in which fertile cells are not protected by a surrounding layer of sterile cells.
4. Cell division in unicellular algae is by membrane infurrowing and cell cleavage. In some green algae, it is by means of a phycoplast. In most multicellular algae, cell division occurs by phragmoplast and outward growth of the cell plate, but in red algae, walls grow inward from the existing side walls.
5. Charophytes have a flagellar root complex of one large band of microtubules, similar to that in plants. Other green algae have a root complex of four bands of microtubules, unlike that of plants.
6. Within green algae, several body types have evolved from unicellular forms: motile and nonmotile colonies, branched and unbranched filaments, coenocytes, membranous forms, and true parenchyma.
7. Several types of life cycle exist: dibiontic in which both a diploid and a haploid generation occurs; monobiontic in which only one generation is capable of mitosis.
8. Dibiontic life cycles may consist of an alternation of either isomorphic or heteromorphic generations. All plants have an alternation of heteromorphic generations.
9. Several evolutionary clades are evident in the green algae, but the charophyte clade is the one that most strongly resembles embryophytes.
10. Brown algae and their relatives make up the heterokont clade, sharing the synapomorphy (derived character) of having one tinsel flagellum and one whiplash flagellum.
IMPORTANT TERMS
agar
alternation of heteromorphic generations
alternation of isomorphic generations
anisogamy
archaeplastid (clade)
autogenous theory
brown algae
charophytes
coccolithophorids
coenocytic (siphonous) body
colony
conceptacles
conjugation
diatoms
dibiontic (species)
dinoflagellates
embryophytes
endosymbiont theory
euglenoids
filamentous body
floridean starch
gametangia
golden-brown algae
green algae
heterokonts
holdfast
isogamous
laminarin
littoral zone
membranous body
monobiontic species
oogamy
oomycetes
parenchymatous body
phycoplast
pit connections
pneumatocyst
primoplantae
receptacles
red algae
red tide
sporangia
streptophytes
trumpet cells
yellow-green algae
zoospores
REVIEW QUESTIONS
1. All organisms are classified into three domains. Name the three and describe the organisms that are classified in each.
2. Reproductive structures are the critical factors in distinguishing algae from true plants of kingdom Plantae. What is one fundamental difference between reproductive structures in these two groups?
3. Which group of algae appears to be most closely related to the ancestors of true plants? Which features appear to be homologous?
4. Describe the endosymbiont theory of plastid origin. How many times might chloroplasts have arisen?
5. In contrast to the endosymbiont theory is the autogenous theory. Describe the basic idea of it. Do you think that the autogenous theory would predict that plastids and mitochondria would have DNA and 70S ribosomes?
6. Why do we think that cyanobacteria rather than purple or green bacteria gave rise to chloroplasts in the primary endosymbiosis? Which cyanobacteria are most similar to chloroplasts? Which algae were involved in the secondary symbioses?
7. Question 6 indicated that chloroplasts arose several distinct times in separate acts of endosymbiosis. If that is true, does the term “plastid” refer to a natural group of organelles that descended from a common ancestor? Would it be safer to use three separate names for these three separate structures?
8. List some of the unusual cyanobacterial features of red algal chloroplasts.
9. Describe cell division and mitosis in algae. How is the red alga method of cross-wall formation different from that of plants?
10. If after cell division the cells adhere to each other loosely, the resulting structure is a ___________________, not an individual organism. In such a structure, are there many types of different cells or are all cells more or less similar to each other?
11. Name and describe the six types of body construction that occur in green algae. What role does the middle lamella play in these? What is the importance of controlling the orientation of cell division? Which of these body types occurs in other groups of algae?
12. Describe monobiontic and dibiontic life cycles. Be careful to mention all possible types. What is the difference between a spore and a gamete? What is the difference between a spore and a zygote?
13. What is an alternation of isomorphic generations? What is an alternation of heteromorphic generations? If you have studied the life cycle of flowering plants, do flowering plants have an alternation of isomorphic or heteromorphic generations?
14. Describe a green alga with a life cycle involving alternation of heteromorphic generations. Why were the two generations originally named as separate species?
15. Volvox is a green alga that you might see in a biology or botany lab. Is it filamentous or membranous or colonial? What is its shape?
16. Spirogyra is another green alga you might see in a biology or botany lab. What is unusual about it? What gives it its name?
17. Chara is an unusual alga. What type of cell division do members of Charophyceae undergo—division with a phycoplast or phragmoplast? What is important about their flagella? Do true plants (especially those with xylem and phloem) have flagella? (Hint: Think sperm cells in ferns and cycads.)
18. Red algae are unusual in many ways, but an especially significant trait is the types of accessory pigments in their chloroplasts. What pigments do they have? They are aggregated into a body called a ___________________.
19. Many red algae have cell walls so heavily impregnated with calcium carbonate that they are brittle. These are called the ___________________ red algae.
20. What are trumpet cells in brown algae? What type of cells do they resemble in true plants? Is their presence in brown algae a sign that brown algae are related to the ancestors of true plants, or is it just a remarkable case of convergent evolution?
21. If you have ever visited a rocky coast, you may have seen two brown algae—kelps and Fucus (these never occur on sandy beaches). Briefly describe the bodies of these two common seaweeds.
22. Dinoflagellates are abundant algae with many unusual features. The arrangement of their flagella is characteristic. Describe the flagella of a typical dinoflagellate.
23. Under ideal conditions, dinoflagellates can reproduce so rapidly that they actually give seawater a distinct color. What is this phenomenon called? Is it safe for fish and humans?
24. Diatoms have what chemical in their cell walls? When the cell dies and the protoplasm degenerates, the wall sinks to the ocean bottom and becomes part of a huge deposit known as ___________________. These deposits can be so extensive because diatoms are so abundant that just a single liter of sea water may easily contain more than ___________________ individuals.
25. Describe and compare the cell walls or cell coverings of dinoflagellates, euglenoids, diatoms, green algae, red algae, and true plants.
26. Describe features of oomycetes that caused people to believe they were fungi (these are believed to be homoplasies). Now describe feature that indicate they are heterokont algae (these are believed to be synapomorphies).
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art