Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Genetics and Evolution
Vascular Plants Without Seeds

Chapter Opener Image: This lower side of a leaf of bird’s nest fern, Asplenium nidus, is covered with rows of sporangia. Within each sporangium, some cells undergo meiosis and produce haploid spores. Just as in bryophytes, these haploid spores are then released and must grow into gametophytes on their own. In seed plants, these spores would be retained, at least temporarily, on the leaf, and would start their development into gametophytes with nourishment from the leaf. In seed plants, megaspores would be retained until they develop fully into seeds; that never happens in the cryptogams (fern, bryophytes, and related plants). The leaf shown here will produce tens of thousands of spores that can be distributed great distances by wind.
OUTLINE
✵ Concepts
✵ Early Vascular Plants
- Rhyniophytes
- Zosterophyllophytes
✵ The Microphyll Line of Evolution: Lycophytes
- Morphology
- Heterospory
- Extant Genera
✵ The Megaphyll Line of Evolution: Euphyllophytes
- Trimerophytes
- Origin of Megaphylls (Euphylls)
- Monilophytes
- Equisetophytes
- Ferns
✵ The Term “Vascular Cryptogams”
Box 21-1 Botany and Beyond: Molecular Studies of the Evolution of Early Land Plants
Box 21-2 Botany and Beyond: Form Genera
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Compare the interpolation hypothesis to the transformation theory.
✵ State the significance of rhyniophytes.
✵ List the characteristics that distinguish zosterophyllophytes from rhyniophytes.
✵ Name the five genera of lycophytes.
✵ Explain how trimerophytes differ from rhyniophytes.
✵ Summarize the telome theory.
✵ Describe the characteristics of ferns.
✵ Explain why the term vascular cryptogams may be used by some to refer to ferns and ferns allies.
 Did You Know?
Did You Know?
✵ Most of the plants in this chapter are ferns.
✵ Ferns never produce flowers, seeds, fruit, or wood.
✵ Some garden ferns act like lilies: During seasons of stress, the foliage dies back and only the underground stems remain alive. They re-sprout when conditions improve.
✵ Ferns are an extremely diverse clade and occupy habitats as different as shady forests, the full sun of deserts, high mountains, and lakes.
![]() Concepts
Concepts
What were the early vascular plants like? What features did they have, and what changes occurred as they diversified and evolved into the many types of plants alive today? Were they simple like some liverworts and hornworts and then gradually became more complex? Or were they completely different, with some giving rise to vascular plants by becoming more complex, whereas others gave rise to nonvascular plants by becoming simpler? Given all of the living plants present now plus the few fossils that we have from the time of the first embryophytes (at least 450 million years ago), what logical hypotheses can be proposed about the early embryophytes?
First, all extant plants (and all known fossil plants) have a dibiontic life cycle: Each species has a multicellular gametophyte and also a multicellular sporophyte. Some algae, especially Coleochaete, are monobiontic, having only one multicellular generation: A zygote undergoes only meiosis, producing more spores that will grow into new gametophytes. The zygote cannot undergo mitosis and cannot grow into a sporophyte. Thus, an important step in the evolution of embryophytes was the conversion of a monobiontic ancestor into dibiontic plants. A sporophyte generation had to come into existence. One hypothesis, the interpolation hypothesis, postulates that a small sporophyte came into existence when a zygote germinated mitotically instead of meiotically (FIGURE 21-1). The sporophyte generation would have gradually evolved in complexity while the gametophyte generation remained small. A sporophyte generation was inserted (interpolated) into the monobiontic life cycle. In this hypothesis, nonvascular plants such as some liverworts might be intermediates in the progression from green algae to vascular plants. For example, the liverworts Riccia and Ricciocarpus have simple, almost algae-like gametophytes, and their sporophytes consist of just a small globose sporangium with no foot or seta. The zygote undergoes several rounds of mitosis, then some cells undergo meiosis. It would take only a few evolutionary modifications to get such a simple sporophyte from an ancestor whose zygote merely underwent meiosis. If so, this sporophyte has been interpolated into the ancestor’s monobiontic life cycle. From a Riccia-like condition, we could hypothesize that the sporophyte would evolve to be more elaborate, with a foot and seta, and then could later evolve to grow by an apical meristem, to branch, to live free of the gametophyte; it could be the ancestor of vascular plant sporophytes. This hypothesis requires that at least one of the nonvascular clades originated very early in the evolution of true plants.
An alternative hypothesis, the transformation theory, postulates that after the dibiontic life cycle originated, both gametophyte and sporophyte became larger, more complex, and vascularized, in a life cycle with an alternation of isomorphic generations (FIGURE 21-2). No living plants have gametophytes that look like sporophytes, but many algae do, and some fossil plants did: Some fossils of early vascular plants bore gametangia and grew among other similar plants that bore sporangia. The transformation theory postulates that these early ancestors diverged into two clades: (1) nonvascular plants in which sporophytes became much simpler and dependent on the gametophytes, and (2) the rest of the vascular plants, in which sporophytes became increasingly elaborate whereas gametophytes became very reduced.
Currently, one issue that is resolved is that all living and most fossil plants are dibiontic with an alternation of heteromorphic sporophytes and gametophytes. What we do not know is whether some liverworts or hornworts are relatively unchanged descendants of very early unvascularized ancestors or whether they originated later from more complex, vascularized ancestors and have since become simpler. This is discussed further after several early vascular plants have been described.
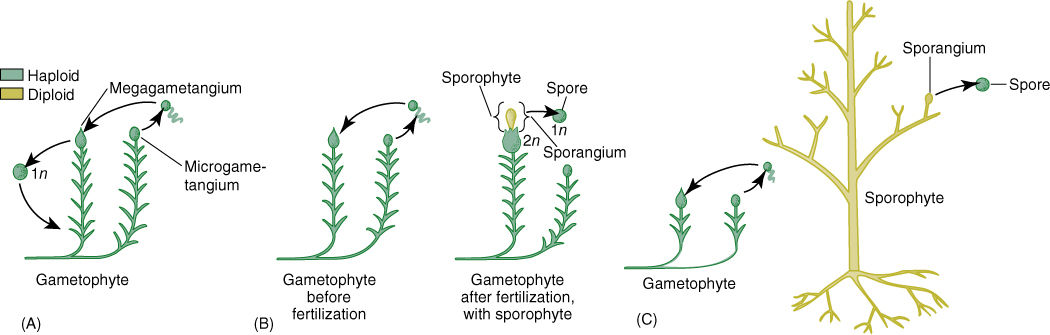
FIGURE 21-1 The interpolation theory. (A) The very earliest land plants were postulated as having no sporophyte; instead, the zygote “germinated” by meiosis, producing haploid spores that grew into new gametophytes. (B) At a later stage in evolution, the zygote would germinate mitotically and produce a simple sporophyte that in the early stages would have consisted of a sporangium and perhaps also a foot; this would have resembled a liverwort sporophyte. (C) With continued evolution, the sporophyte would have become progressively more elaborate while the gametophytes became simpler.
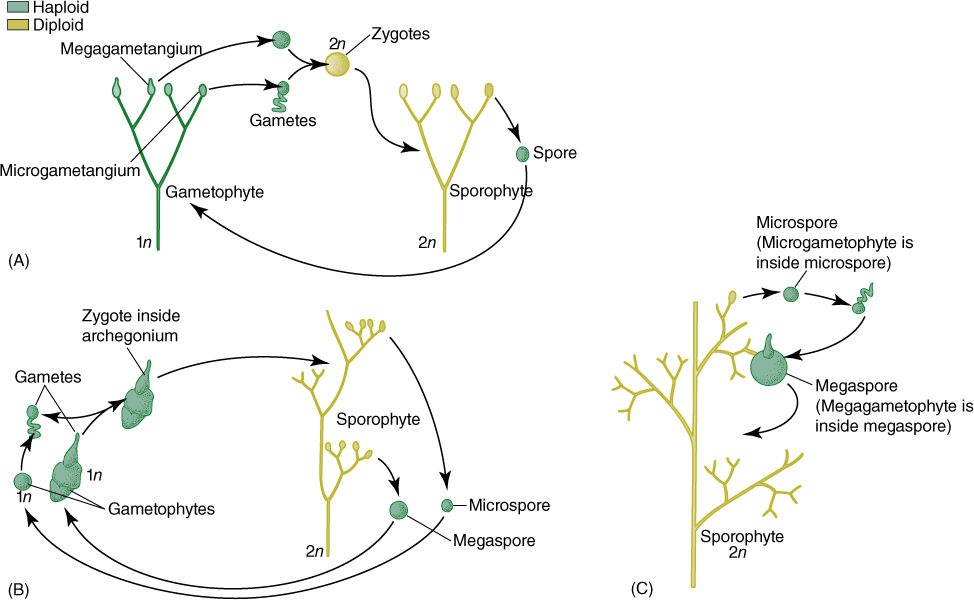
FIGURE 21-2 The transformation theory of the origin of the vascular plant life cycle postulates that in early land plants (A), gametophytes were upright and dichotomously branched, with epidermis, cuticle, and vascular tissue, just like sporophytes. (B) With time, sporophytes became larger and more complex, and gametophytes became simpler. In the species illustrated here, gametophytes have become so small that the microgametophyte develops within the spore wall and the megagametophyte protrudes from the spore only slightly. (C) With continued reduction, it is possible—but neither necessary nor inevitable—for the megaspore and its megagametophyte to be retained inside the megasporangium and remain on the parental sporophyte, an important step in the process of seed evolution.
![]() Early Vascular Plants
Early Vascular Plants
Rhyniophytes
The earliest fossils that definitely were vascular land plants belong to Cooksonia, a genus of extinct plants (FIGURE 21-3). These had upright stems that were simple, short cylinders (several centimeters long) with no leaves (they had “naked stems”). They had equal dichotomous branching, both branches being of equal size and vigor (FIGURE 21-4). Plants of Cooksonia had an epidermis with a cuticle, a cortex of parenchyma, and a simple bundle of xylem composed of tracheids with annular secondary walls. The ends of the branches were swollen and contained large, multicellular masses of sporogenous tissue surrounded by several layers of sterile cells. As in all plants, only the central cells were sporogenous, and the sporangium had to open to release the spores. Plants of Cooksonia were homosporous; there were no separate microspores and megaspores. Fossils that have these general characters are called rhyniophytes.
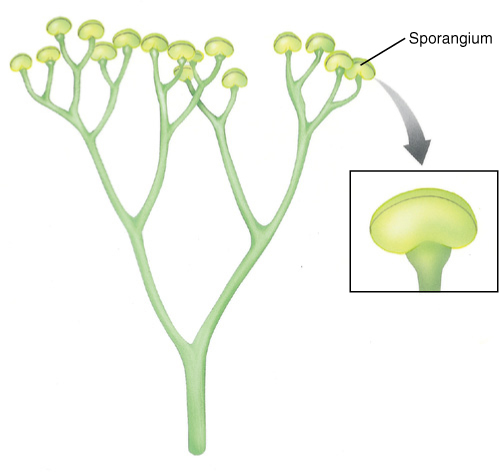
FIGURE 21-3 A reconstruction of Cooksonia caledonica, the earliest known plant that had xylem—tracheids with annular secondary walls. Important features are its sporangia at the ends of branches, the lack of leaves, and the dichotomous branching.
Rhynia and Aglaophyton were other rhyniophytes, early vascular plants similar to Cooksonia (FIGURE 21-5). They had a prostrate rhizome, upright naked stems, and terminal sporangia. Stomata and guard cells occurred in the epidermis, and there was a layer that appears to have been a cuticle.
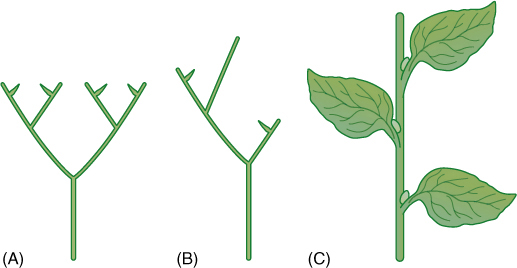
FIGURE 21-4 Branching patterns. (A) If a stem forks, resulting in two equal stems, it is a dichotomy or dichotomous branching. (B) It is pseudomonopodial branching if one stem is definitely larger and tends to form a trunk. (C) If one stem dominates the system absolutely, it is monopodial branching, as in most seed plants.
In the same rocks with Rhynia and Aglaophyton are fossils of similar plants. Two, Lyonophyton and Sciadophyton, were gametophytes, not sporophytes. The ends of their stems bore flattened cup-shaped areas that contained gametangia, both antheridia and archegonia, but not sporangia. These plants had upright, dichotomously branched stems, vascular tissue, stomata, and a cuticle. These were much larger and more complex than the gametophytes of any mosses, liverworts, or hornworts that are alive today. It is possible to suspect that because Rhynia-type sporophytes occur together with Sciadophyton-type gametophytes, they might have been alternate phases of the same species. If true, these plants had an alternation of isomorphic generations (Figure 21-2A), and later evolution into seed plants involved reduction of the gametophyte to just a few cells and elaboration of the sporophyte into a more complex plant. These may have given rise to the nonvascular plants by reduction of both gametophytes and sporophytes.
Another fossil from that same time, Horneophyton, had sporophytes with naked axes that branched dichotomously and were up to 20 cm tall (Figure 21-5C). They had stomata and terminal sporangia. Each sporangium had a short columella, and just as in hornworts, the sporogenous tissue surrounded the columella on all sides and the top. The stem base was swollen, similar to a hornwort foot, except that it bore rhizoids. The similarities between the hornworts Anthoceros and Phaeoceros and Horneophyton raise the possibility that vascular plants may have arisen from hornwort-like ancestors (Figure 21-5D, from left to right), but it is also possible to think about these transitions in reverse—that hornworts evolved from species like Horneophyton by means of an elaboration of the gametophyte and a reduction of the sporophyte (Figure 21-5D, from right to left).
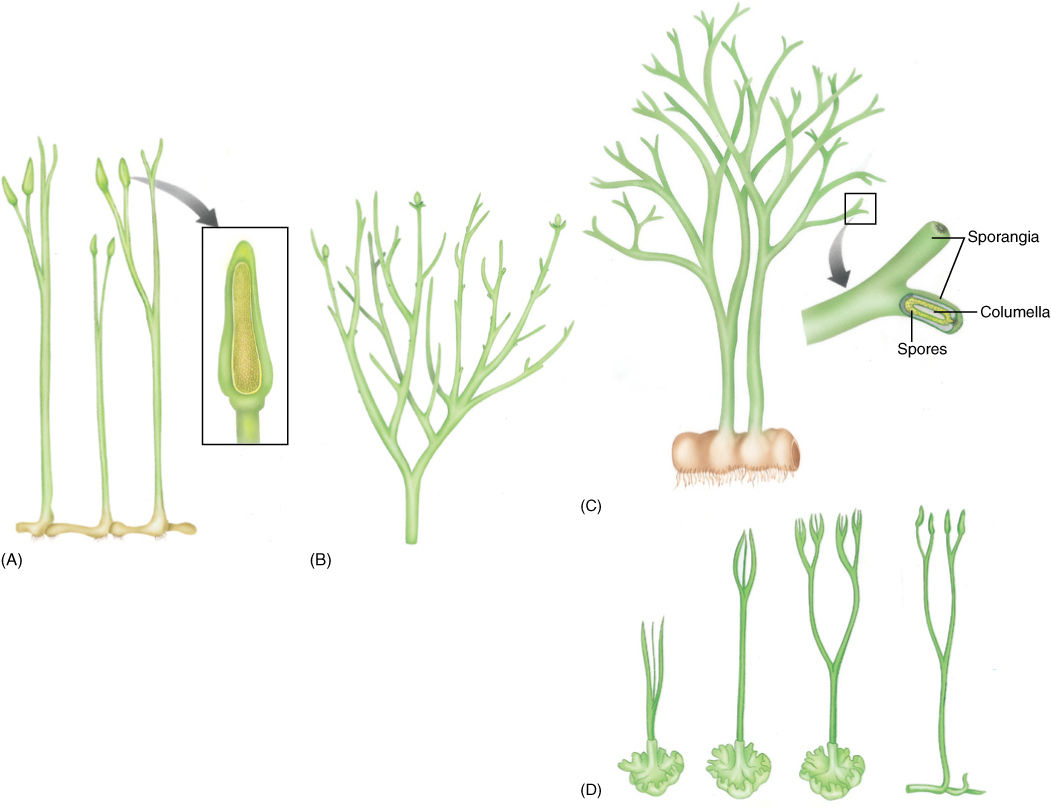
FIGURE 21-5 Reconstruction of Aglaophyton major (previously known as Rhynia major) (A) and Rhynia gwynne-vaughanii (B). They strongly resemble Cooksonia, and we know that they definitely had rhizomes, upright stems, and rhizoids. Inset shows the sporangium cut away, revealing spores. (C) Reconstruction of the extinct plant Horneophyton and its terminal sporangia. (D) At first, it was postulated that algae gave rise to true plants whose life cycle was dominated by the gametophyte and whose sporophyte was small and dependent, hemiparasitic on the gametophyte (as drawn on the left). It was postulated that this evolved into the vascular plants as the sporophyte became larger and more elaborate while the gametophyte became reduced (as drawn on the right). But it is possible that hornworts evolved by the reverse process, reading this series from right to left.
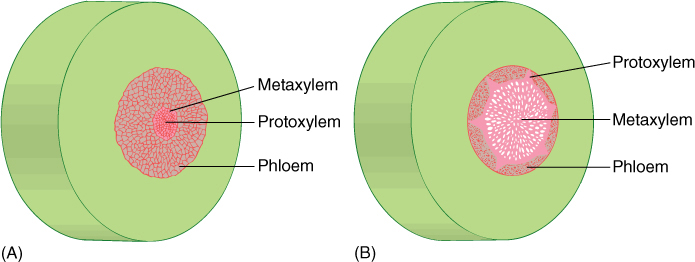
FIGURE 21-6 Early vascular plants had two types of organization of xylem in their stems. In both, the center is a solid mass of xylem with no pith. This is a protostele. (A) In an endarch protostele, protoxylem is located in the center; metaxylem is on the periphery. Endarch protosteles occurred in the rhyniophytes. (B) In an exarch protostele, metaxylem is located interior to the xylem mass and protoxylem is on the edges, as several groups. Exarch protosteles are found in fossils of zosterophyllophyte stems.
Xylem Structure of Early Vascular Plants
Early vascular plants had two types of xylem organization. In both, the center is a solid mass of xylem with no pith; this is a protostele. In an endarch protostele, protoxylem is located in the center and metaxylem differentiates on the outer edge of the xylem mass (FIGURE 21-6A). Protoxylem is the xylem that differentiates while cells are small and narrow, and metaxylem differentiates after the cells have expanded for a few more hours or days and are larger. The other type of stele present in early vascular plants is an exarch protostele, with metaxylem located in the center of the xylem mass and protoxylem on the edges as several groups next to the phloem (FIGURE 21-6B). Another type of stele, which did not evolve until later, is the siphonostele, one in which pith is present in the center, as occurs in the stems of ferns and seed plants (see Figure 21-29). Because xylem is often preserved well in fossils, its characteristics of exarch/endarch and protostele/siphonostele are usually available for study.
The xylem in many specimens of Rhynia and Aglaophyton is well preserved: It was a round cylinder without pith but with protoxylem in the center and metaxylem on the exterior—that is, an endarch protostele. All xylem cells were tracheids with annular thickenings. Around the xylem was a layer of phloem-like cells and then a parenchymatous cortex and epidermis.
Zosterophyllophytes
Another group of early vascular plants are the zosterophyllophytes, named after the principal genus Zosterophyllum (FIGURE 21-7A). They were small herbs without secondary growth. Many of their features were similar to those of rhyniophytes, but three characteristics make us think they were a distinct group: Their sporangia were lateral, not terminal; sporangia opened transversely along the top edge (FIGURE 21-7B), and their xylem was an exarch protostele, that is, protoxylem on the outer margin and metaxylem in the center (see Figure 21-6 and TABLE 21-1).
Although these distinctions may seem minor, we are certain that in all the more recent plants with large leaves (the euphyllophytes: ferns, conifers, flowering plants), sporangia are terminal rather than lateral; thus, plants such as Rhynia may have been the transitions between algae and later seed plants, but Zosterophyllum was not. Instead, some of the simplest vascular plants alive today, the lycophytes, have lateral sporangia, and they may represent a line of evolution based on Zosterophyllum-like ancestors (FIGURE 21-8).
Zosterophyllum plants (all are extinct) grew as small bunches, only approximately 15 cm high. Upper portions of their stems had cuticle, ordinary epidermal cells, and stomata, but lower portions did not, presumably because they grew in swampy, marshy areas. Stems of Zosterophyllum were naked (smooth), branched dichotomously, and contained a small amount of xylem that consisted of tracheids with annular and scalariform secondary walls. Sporangial walls were several layers thick, and all spores were the same size; therefore, they must have been homosporous.
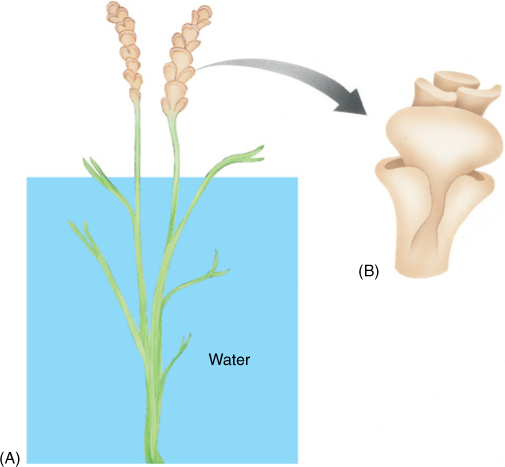
FIGURE 21-7 (A) Zosterophyllum rehenanum plants were quite similar to those of rhyniophytes, but the ends of fertile branches bore numerous lateral sporangia, not a single terminal one. Also, sporangia (B) opened by a suture that passed over the top of the sporangium, not up its side.
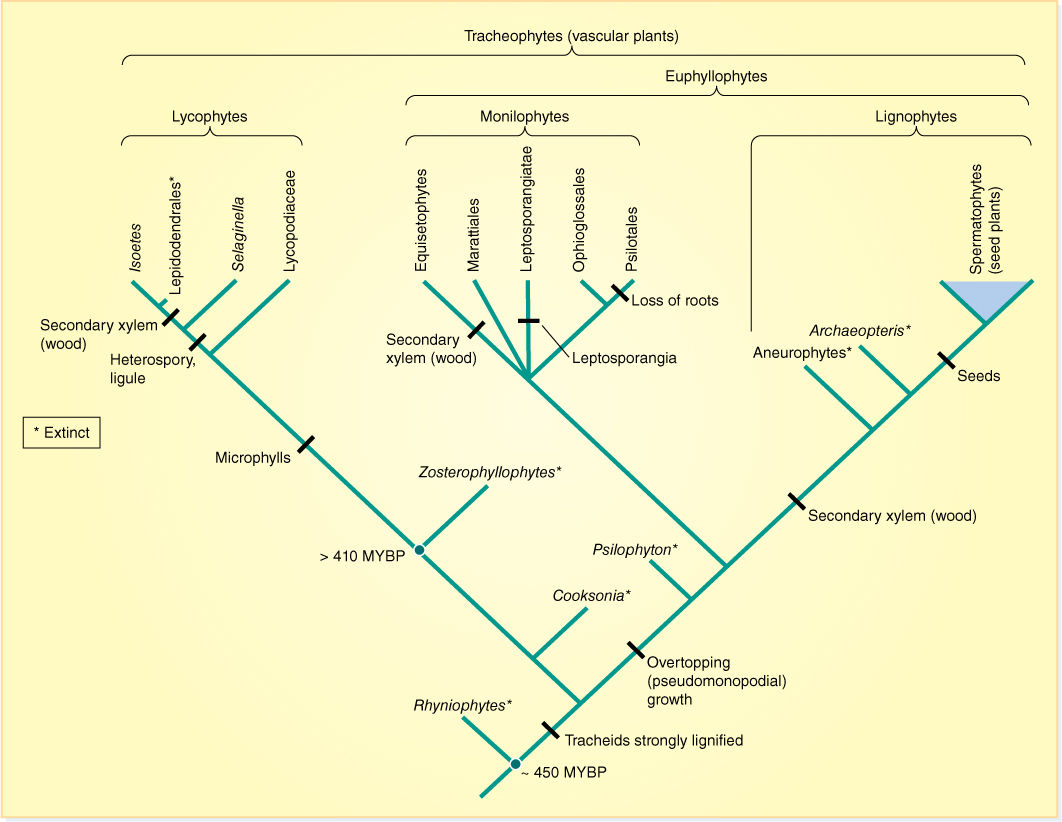
FIGURE 21-8 One proposed phylogeny of vascular plants without seeds (MYBP = million years before present).
TABLE 21-1 Characteristics of Rhyniophytes and Zosterophyllophytes
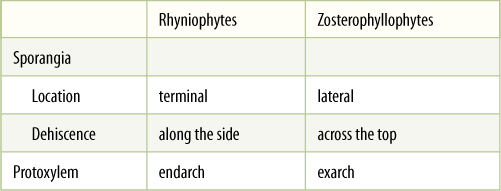

FIGURE 21-9 This reconstruction of the zosterophyllophyte Rebuchia ovata shows several significant advances in body construction. The stems that bore sporangia were specialized, keeping sporangia close together and elevated into the wind; other stems branched repeatedly, forming a large photosynthetic surface. Some branches were larger than others, and thus, branching was no longer strictly equal dichotomies.
Other genera of zosterophyllophytes show that significant morphological changes evolved quickly. In Rebuchia, sporangia occurred together on the ends of specialized branches (FIGURE 21-9). In Crenaticaulis, some branching was pseudomonopodial: Larger, trunk-like shoots bore smaller, shorter lateral shoots. These are the morphological bases for producing cones and trunks. Several zosterophyllophytes had a smooth surface (see Figure 21-7), but others had outgrowths called enations that ranged from quite small to long, thin scales. Enations increased the photosynthetic surface area of the plants, and in Asteroxylon, they contained stomata and a small trace of vascular tissue that ran from the stele through the cortex to the base of the enation (FIGURE 21-10).
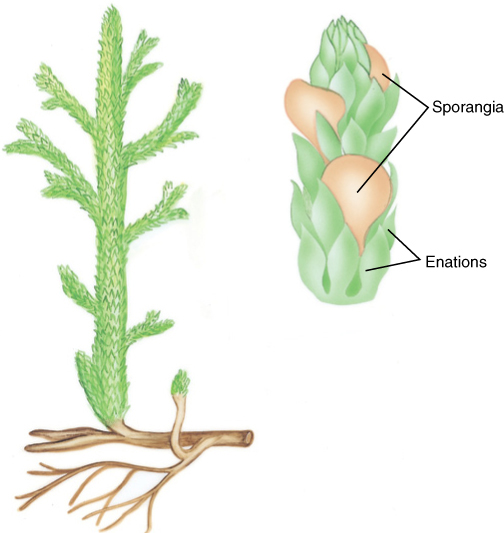
FIGURE 21-10 Reconstruction of Asteroxylon, an early lycophyte, showing its surface covered with enations, small flaps of photosynthetic tissue. In species without enations, stems were round and oriented vertically, not very good for harvesting sunlight.
![]() The Microphyll Line of Evolution: Lycophytes
The Microphyll Line of Evolution: Lycophytes
Lycophytes represent a distinct line of evolution out of the early land plants that resembled zosterophyllophytes. Lycophytes have lateral sporangia and exarch protosteles, and thus, they may have come from a Zosterophyllum type of ancestor (Figure 21-8 and TABLE 21-2).
Morphology
The earliest lycophytes were members of the genera Drepanophycus and Baragwanathia (FIGURE 21-11A). They were similar to their presumed ancestors, the zosterophyllophytes, with an important difference: Their enations were large, up to 4 cm long, and they contained a single well-developed trace of vascular tissue. Such enations must have increased photosynthesis, and they could be called leaves; however, “leaf” is an ambiguous term, and enations in the division Lycophyta are called microphylls for clarity (FIGURES 21-11B—E; the term “lycophyll” has been suggested recently). “Micro-” refers to their evolution from small enations, not to their actual size. In some plants, they were up to 78 cm long. This is not the line of evolution that led to ferns and seed plants; microphylls are not the same as the leaves you are familiar with.
TABLE 21-2 Classification of Division Lycophyta
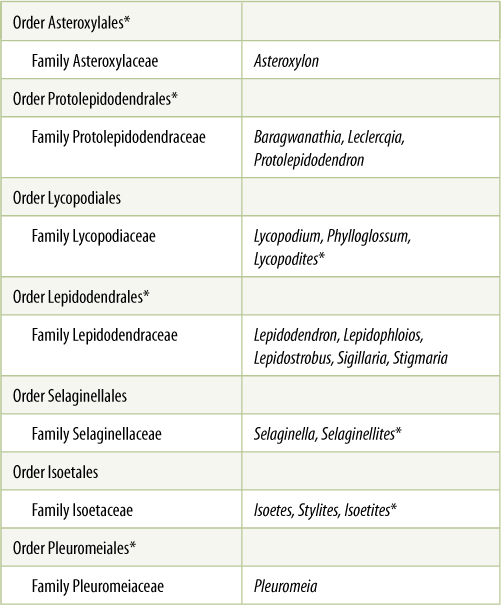
*All species in this group are extinct.
Another important advance was the evolution of true roots that allowed lycophyte sporophytes to anchor firmly, absorb efficiently, and thus to grow to tremendous size.
Many extinct lycophytes such as Lepidodendron, Sigillaria, and Stigmaria had a vascular cambium and secondary growth (FIGURES 21-12 to 21-14). Their wood looked remarkably like the secondary xylem of pines and other living conifers, having a pith, rays, and elongate tracheids. However, the vascular cambium had one major flaw: Its cells apparently could not undergo radial longitudinal division; therefore, new fusiform initials could not be produced. As the wood grew to a larger circumference, cambial cells became increasingly wider tangentially (Figure 21-14B). No fossil has ever been found with wood more than about 10 cm thick. After that much secondary growth, the cambial cells may have stretched so much that they could no longer function.
Heterospory
In many extinct and extant lycophytes sporangia are clustered together in compact groups called cones or strobili (singular, strobilus), which protect them (FIGURE 21-15). Although many species remained homosporous, others became heterosporous, having microspores and megaspores that germinated to give rise to distinct microgametophytes and megagametophytes, respectively. Heterospory is a necessary precondition for the evolution of seeds (see Figure 21-2). In the lycophytes Lepidostrobus and Lepidophloios, the megaspore developed into a megagametophyte without enlarging: The megagametophyte existed completely within the wall of the original megaspore, which was up to 10 mm long. Furthermore, the megaspore in some species was retained within the sporophyll, protected by thick-walled cells of the sporangium. This is remarkably similar to ovules and seeds in seed plants, the most important difference being that in these lycophytes, the sporangium dehisced (much like modern anthers do), and the megaspore wall cracked, exposing the archegonia. Sperm cells could swim to the egg during fertilization; pollen tubes were unnecessary.
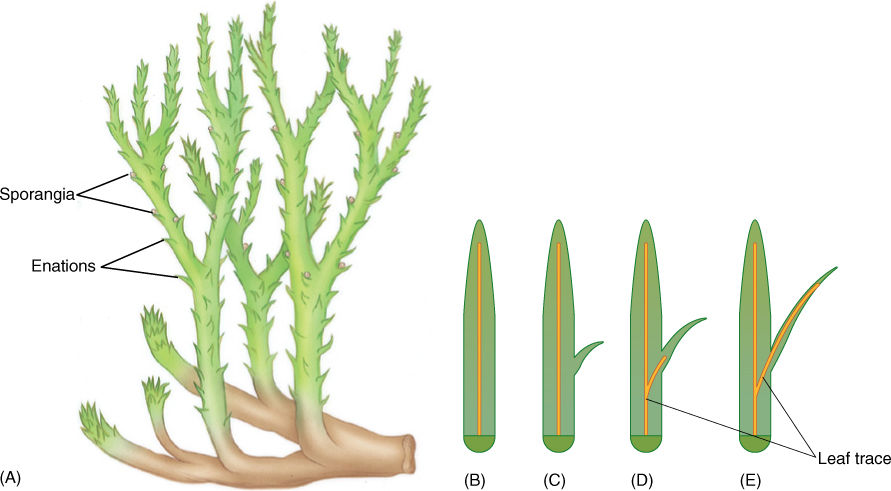
FIGURE 21-11 (A) Drepanophycus was an early lycophyte that was still small and simple. (B) — (E) Microphylls in lycophytes are believed to have evolved as enations. Originally they were small, simple flaps of photosynthetic tissue (C). Later they became larger (D) and were vascularized (E). See Figure 21-20 for evolution of megaphylls.
Lycophytes are remarkable in that they represent an ancient line of evolution distinct from seed plants but having convergent evolution in several characters: leaves, roots, secondary growth, and almost seeds. In the Devonian and Carboniferous Periods (see diagram on the inside of the back cover), this group dominated the swampy areas of Earth with extensive forests of large trees (Figure 21-12), but most became extinct; currently, the entire division contains only five genera, Lycopodium, Phylloglossum, Selaginella, Isoetes, and Stylites.

FIGURE 21-12 During the Carboniferous Period, Earth’s vegetation was dominated by extensive forests of large lycophyte trees. None became truly massive like our redwoods, oaks, and elms, but the forests were both dense and vertically stratified. Flying insects were present, and early reptiles were just appearing; amphibians were the dominant land animals.
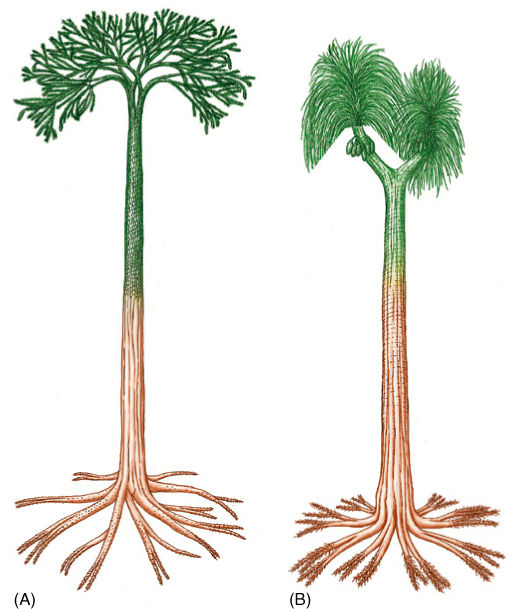
FIGURE 21-13 (A) Reconstruction of Lepidodendron showing a large tree with one distinct trunk (monopodial growth) and a well-developed root system. Leaves (microphylls) were sophisticated, and sporangia occurred in discrete strobili (cones). (B) Sigillaria was a large lycophyte tree in which the leaves usually were 1 m or more long. On the branch to the left, leaves have been left undrawn to reveal cones of sporangia.
Extant Genera
Lycopodium (“ground pine” or “club moss”) is fairly common in forests from tropical regions to the arctic (Figure 21-15). All living species, approximately 200, are small herbs with prostrate rhizomes that have true roots and short upright branches. Microphylls are spirally arranged on their stems, and secondary growth never occurs. Sporangia may be arranged in cones or distributed along the shoots (L. lucidulum). All Lycopodium species are homosporous, a plesiomorphic (relictual) trait. Spores germinate and grow into bisexual gametophytes that produce both antheridia and archegonia. In some species, gametophytes are green and photosynthetic; in others, they are subterranean and heterotrophic, nourished by fungi.
Selaginella is less common in temperate North America, and its plants are smaller and easily overlooked or mistaken to be mosses (FIGURE 21-16). Probably the best known species is the resurrection plant, S. lepidophylla, which curls up, turns brown, and appears dead upon drying but uncurls and regreens when moistened. Unlike Lycopodium, Selaginella has the apomorphic (derived) condition of being heterosporous, and the megagametophyte develops inside the megaspore wall (FIGURE 21-17A). The megaspore is not retained on the sporophyte, however, and is not seed like. Microgametophytes also develop within the spore wall and consist of a single vegetative cell and an antheridium (FIGURE 21-17B). Many flagellate sperms are produced and then released as the spore wall ruptures.
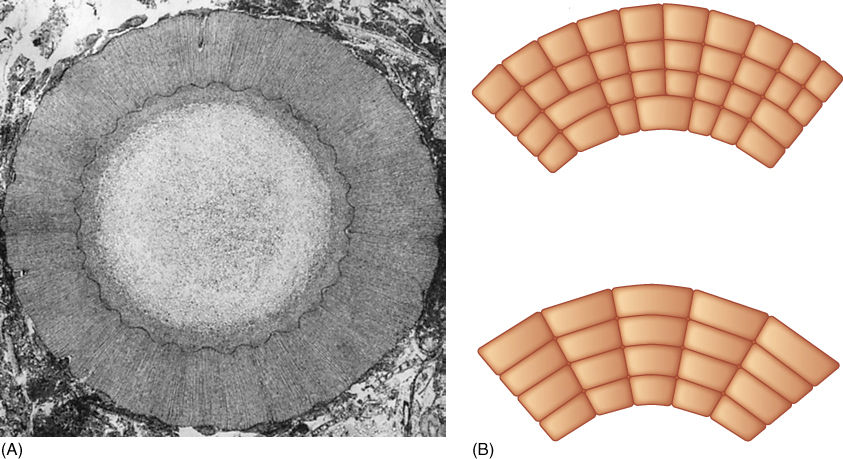
FIGURE 21-14 (A) Wood of Sigillaria consisted of large tracheids and superficially resembled the wood of modern conifers; however, it was not as derived in its pitting and other features of its secondary wall, and it was formed by a vascular cambium that could not form new fusiform initials (× 80). (B) In the vascular cambia of seed plants (top), as the cambium is pushed outward by accumulation of new secondary xylem, its circumference becomes greater, but fusiform initials divide by radial walls, creating new fusiform initials. An old cambium has many more cells than a young cambium. In arborescent lycophytes (bottom), cambial cells could not undergo this division; therefore, an old cambium had no more cells than a young one. Older cells became stretched circumferentially until they were no longer functional; secondary growth then ceased.

FIGURE 21-15 (A) Lycopodium cernuum is a common species that has extensive rhizomes, vertical chlorophyllous shoots, and sporangia clustered into cones. These shoots are leaning against and being supported by surrounding grasses and shrubs. (B) Lycopodium obscurum, with sporangia clustered into cones at the tips of branches. (C) and (D) Lycopodium lucidulum is one of several lycopod species in which sporangia are distributed among the leaves rather than in strobili.
Selaginellas can be distinguished from lycopodiums by a small flap of tissue, the ligule, on the upper surface of Selaginella leaves. Although ligules are simple structures, and their adaptive advantages are unknown; they are ancient and can be used to distinguish early fossil selaginellas from fossil lycopods.
Isoetes is a genus of about 60 species of small, unusual plants called quillworts, which grow in wet, muddy areas that occasionally become dry (FIGURE 21-18). Their body consists of a small corm-like stem that has roots attached below and leaves above. Isoetes is heterosporous like Selaginella, and almost every leaf contains sporangia. Microphylls in this genus also have ligules. Weak cambial growth results in the production of additional cortex parenchyma to the exterior and a type of vascular tissue to the interior; the latter tissue is a mixture of tracheids, sieve elements, and parenchyma. Stylites plants are very similar to those of Isoetes, and several botanists have suggested that the two species of Stylites are really extreme forms of Isoetes.
The anatomy and morphology preserved in fossil lycophytes indicate many instances of convergent evolution with seed plants. In both lines, elaborate, efficient leaves, wood, bark, and roots evolved. Sporangia became separated from nonreproductive organs and were grouped together into strobili. In both lycophytes and seed plants, heterospory evolved, as did endosporial development of the megagametophyte. In seed plants, megaspores were retained in the mega-sporangium and evolved into seeds; this nearly happened in certain extinct lycophytes. We cannot tell from the fossils whether convergent evolution of specialized metabolisms also occurred, but Isoetes does have CAM (Crassulacean acid metabolism) photosynthesis. It also has a unique means of obtaining its carbon dioxide—absorption by the roots from soil or mud. Its leaves have a thick cuticle but no stomata. We can only wonder what the metabolism of the extinct tree-like lycophytes was like. The extinction of so many of the lycophytes is unfortunate; they were complex, sophisticated plants that had many superb adaptations. They probably underwent interesting responses to changes in season, and they must have been able to resist numerous types of pathogenic bacteria, fungi, and insects. Those aspects of their biology will probably remain unknown to us forever.

FIGURE 21-16 (A) Plants of Selaginella willdenowii are large and occur in moist habitats. (B) Plants of S. lepidophylla are also large, but occur in habitats that cycle between wet and severely dry. When dry, as shown here, the plant desiccates, curls up, and becomes dormant; the brown color is only on the underside of its leaves—the upper sides are green, even in this dry condition. If moistened with rain or dew, it quickly rehydrates, uncurls, and becomes active. (C) Plants of S. wrightii tend to be small, compact, and easily mistaken for mosses. These are desert adapted and grow on bare rock in full sun throughout arid regions of the western United States.
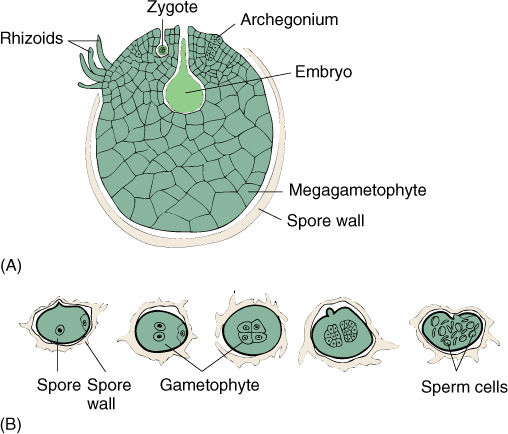
FIGURE 21-17 (A) The megagametophyte of some selaginellas develops almost entirely within the megaspore wall. The spore cracks open, exposing archegonia (megagametangia) and permitting fertilization by swimming sperm cells. (B) Microgametophytes of selaginellas develop within the microspore wall, then liberate motile sperms.

FIGURE 21-18 Although Isoetes is a small plant, its short basal corm-like stem has a small amount of secondary growth.
![]() The Megaphyll Line of Evolution: Euphyllophytes
The Megaphyll Line of Evolution: Euphyllophytes
Trimerophytes
Division Trimerophytophyta was proposed in 1968 for three genera of extinct plants, Trimerophyton, Psilophyton, and Pertica. Their fossils strongly resemble those of rhyniophytes, having terminal sporangia that dehisced laterally, homospory, dichotomous branching, and an endarch vascular cylinder of tracheids. Trimerophytes, however, are considered a distinct advancement out of rhyniophytes because of several special features. Most important is the trend of overtopping: Trimerophytes had an unequal branching in which one stem was more vigorous (see Figure 21-4B). In later species, the inequality was so pronounced that main stems and lateral stems can be identified easily. Finally, Pertica displays pseudomonopodial branching, that is, a single main trunk rather than a series of dichotomies (FIGURE 21-19). The plants have small lateral branches, some fertile and bearing sporangia and others sterile and acting as leaves.
Simultaneously, the positioning of branches became more regular and controlled. In rhyniophytes, the points of dichotomy were irregular and unpredictable, but in some species of trimerophytes, lateral branches were arranged in a regular spiral phyllotaxy. Other types of phyllotaxy that occurred were alternate, decussate (opposite leaves arranged in four rows), distichous (leaves in two rows), tetrastichous (alternate leaves, in four rows), and even whorled. Correlated with the evolution of a pseudomonopodial growth habit and the presence of numerous photosynthetic lateral branches was an increase in the vigor and robustness of the plants. Individuals of Pertica quadrifaria had stems 1.5 cm wide and about 1 m tall, whereas those of P. dalhousii are estimated to have been as much as 3 m tall, approximately the height of tall shrubs today. Although a dense stand of P. dalhousii would not be called a forest, there would have been vertical stratification of light, air movement, and humidity within the canopy.
Trimerophytes became distinct from rhyniophytes during the Lower Devonian and existed until the Upper Devonian Period and then came to an end not by going extinct but rather by evolving into the ancestors of ferns and seed plants.
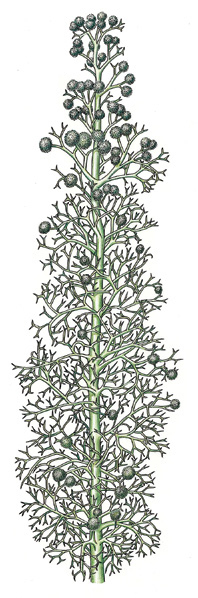
FIGURE 21-19 Pertica quadrifaria had one main trunk from which grew small branches; the smallest twigs still show dichotomous branching, but larger stems branched pseudomonopodially. This branch pattern also evolved separately in lycophytes, but here in the trimerophytes, it is the ancestor to the stem structure of seed plants. The globular structures are clusters of sporangia.
Origin of Megaphylls (Euphylls)
At least three distinct types of homoplasic (analogous) structures called leaves occur in plants: (1) leaves on gametophytes of nonvascular plants; (2) enations/microphylls of zosterophyllophytes and lycophytes; and (3) megaphylls, leaves that evolved from branch systems and are present in all seed plants, ferns, and equisetophytes. Megaphyll evolution is summarized by the telome theory. Imagine a plant like Pertica consisting of a main stem and smooth, cylindrical, dichotomously branching lateral stems (Figure 21-19). The ultimate twigs, those of the last dichotomy, are known as telomes. Now imagine that all subdivisions of a lateral branch become aligned in one plane (planation; FIGURE 21-20) and that parenchyma develops between telomes and even lower branches (webbing). This is not a leaf, but it has suitable characteristics to be the ancestor to leaves. Also, the leaves we see on trees and herbs of angiosperms are the result of about 300 million years of evolutionary refinement. If the branch system involved in this evolution produced sporangia, the resulting structure would not be just a leaf, but rather a sporophyll, a sporangium-bearing, leaf-like structure. For one reason or another, the plants with this organization outcompeted those with microphylls, the lycophytes. This is not to say that the megaphyll type of organization itself was the main reason for their success, but today, megaphyllous plants are by far the more common. We believe that all megaphyllous plants form a monophyletic clade, now referred to as the euphyllophytes.
Monilophytes
Several current studies suggest that megaphyllous plants (euphyllophytes) are united by three synapomorphies: (1) their roots have exarch xylem, (2) they have megaphylls, and (3) they have a 30-kilobase inversion in the large single-copy region of their plastid DNA (BOX 21-1). The euphyllophytes as presented in Figure 21-8 contains two sister clades, the monilophytes and the woody plants (lignophytes). Many monilophytes are plants we know as ferns (Leptosporangiatae, Marattiales, Ophioglossales); others have been called “fern allies” (equisetophytes, Psilotales). If we want to use the term “ferns” in the sense of a monophyletic, natural group, we must also refer to equisetophytes (horsetails) and Psilotales as ferns. This sounds strange to many experienced botanists because horsetails differ from all other ferns in many respects. Remember that Figure 21-8 is a model, a hypothesis. It would not be surprising to some of us if within a few years, new data would indicate that equisetophytes form a clade sister to ferns, but not part of ferns themselves. In this book, Equisetum and its clade will be treated as sister to the ferns.
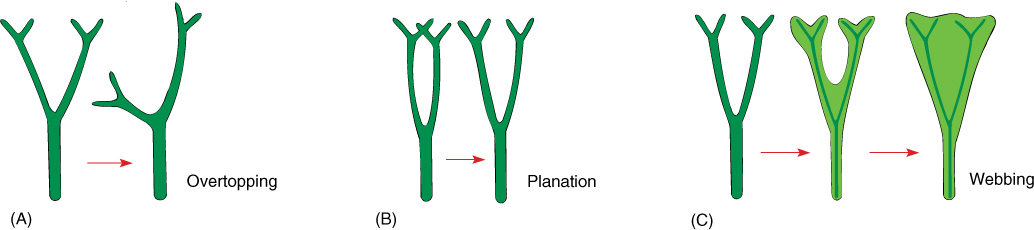
FIGURE 21-20 The leaves of ferns, Equisetum, and seed plants are megaphylls (euphylls) that arose by evolutionary modification of stems. (A) In the first step, one branch (the main axis) overtops the other, which remains smaller and lateral. (B) Next, lateral branch systems restricted their branching to just one plane (planation); they stopped producing three-dimensional branch systems. (C) Finally, the spaces between close branches developed a thin sheet of chlorophyll-containing cells in a type of webbing.
Equisetophytes
Equisetophytes have been classified as division Arthrophyta (also called Sphenophyta). They consist of several genera of extinct plants and one genus, Equisetum, with 15 extant species known as horsetails or scouring rushes (TABLE 21-3). The living plants are all herbs without any secondary growth, and although certain species may attain a height of up to 10 m, they are usually less than 1 m tall (FIGURE 21-21). Their aerial stems have a characteristic jointed structure, with a whorl of fused leaves at the nodes. The leaves are small and have just a single trace of vascular tissue, but they are small megaphylls, not microphylls. If branches are present, they alternate with the leaves rather than being located in the leaf axils. The vertical, aerial stems arise from deep subterranean rhizomes; in harsh habitats, aerial shoots die during winter, but rhizomes persist and plants can spread vigorously. True roots are present, being produced at the rhizome’s nodes.
TABLE 21-3 Classification of Division Arthrophyta
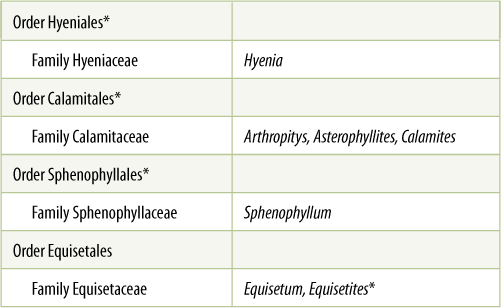
*All species in this group are extinct.

FIGURE 21-21 Equisetum plants are usually small, having vertical shoots that arise from subterranean, highly branched rhizomes. In some species such as E. hyemale (A), each shoot is both photosynthetic and reproductive; in other species, such as E. arvense (B), some shoots are vegetative only (shown here), and separate reproductive shoots are produced at a different time of year. (C) This is E. giganteum, in which the plants grow exceptionally large.
Botany and Beyond
BOX 21-1 Molecular Studies of the Evolution of Early Land Plants
The evolutionary relationships of early land plants are still not understood well. Plants like mosses, liverworts, Rhynia, and lycopods are so simple that there are not many characters to compare. Furthermore, when two groups do resemble each other, we cannot always be certain whether the common characters are homologous (derived from the same ancestral character) or analogous (derived from different ancestral characters and resembling each other only as a result of convergent evolution).
Analysis of the arrangement of genes in chloroplasts has been used to investigate which early land plants are related. The circular DNA from chloroplasts has been isolated from many plants; then DNA restriction enzymes were used to analyze the order in which genes occur. Currently, many groups have been studied, and it turns out that mosses, liverworts, and lycophytes have one arrangement whereas euphyllophytes all have an alternative arrangement.
From numerous studies on many types of plants, it had been concluded that chloroplast genomes are quite stable and this type of rearrangement is rare. It seems unlikely that this type of rearrangement could occur twice, producing similar gene arrangements in three distinct groups. Thus, the presence of the rearrangement in all euphyllophytes is strong evidence that all constitute a monophyletic clade, that monilophytes, early woody plants, and modern seed plants all shared a common ancestor.
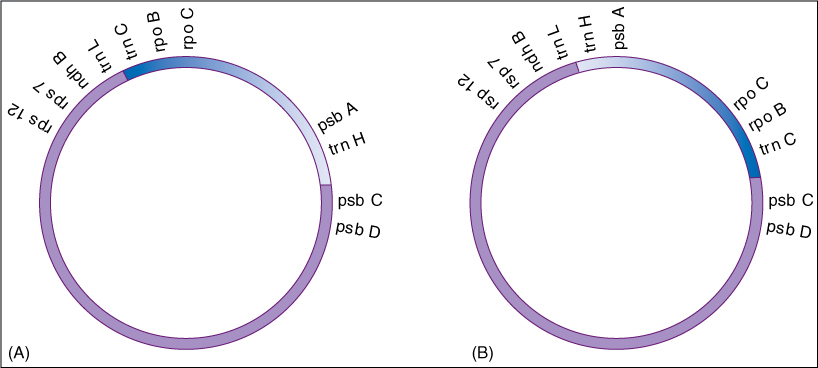
FIGURE B21-1 The order of genes on the circular DNA of plastids of liverworts, mosses, and lycophytes (A), and of seed plants (B). The section shown in blue is reversed in euphyllophytes.
Internal structure is as distinctive as external morphology (FIGURE 21-22). Stems have a pith, and thus, these are siphonosteles, not protosteles as in all other plants mentioned so far (compare Figure 21-22 with Figure 21-6). Protoxylem forms next to the pith, interior to the metaxylem, and thus, it is endarch, a rhyniophyte/trimerophyte trait distinctly different from the exarch protoxylem of zosterophyllophytes. Vessels are rare outside the flowering plants, occurring only in Selaginella, several ferns, three genera of gymnosperms, and Equisetum. Stem elongation causes some cells to be stretched and torn, forming canals. The cortex of Equisetum is composed of large cells, the outer ones chlorophyllous. Air canals may be located in the outer cortex. The epidermis contains stomatal pores and guard cells, as well as such large amounts of silica that the stems are tough and rigid.
Reproductive structures in Equisetum are specialized; sporangia always occur in groups of five to ten located on an umbrella-shaped sporangiophore (FIGURE 21-23). This has a short stalk and a flat, shield-shaped head from which the sporangia project, parallel to the sporangiophore stalk. Sporangiophores are always arranged in compact spirals forming a strobilus. Equisetum is homosporous; therefore, all plants have only one type of sporangiophore and strobilus, but in some species, the strobilus occurs at the tip of a green photosynthetic shoot, whereas in others it is borne on a special, colorless reproductive shoot.
After spores are released, they germinate on moist soil and develop into small (1 mm across) green gametophytes. They have no vascular tissue and no epidermis or stomata; the body is just a small mass of parenchyma. Gametophytes of Equisetum are either male or bisexual. Antheridia release numerous multiflagellate sperms that swim to the archegonia and eggs. The megagamete and zygote are never released, but rather are retained and nourished by the gametophyte.
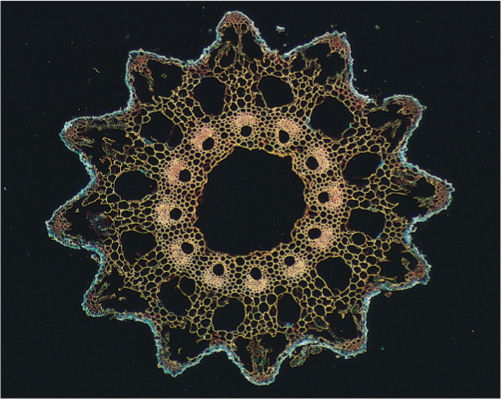
FIGURE 21-22 The stem anatomy of Equisetum is dominated by numerous canals. The center of the pith (this is a siphonostele) is torn apart, forming a central canal or pith canal, and around the remnants of the pith are carinal canals formed by breakdown of protoxylem. Just exterior to the carinal canals are metaxylem and phloem; vascular bundles are endarch. The cortex contains cortical canals that alternate with carinal canals, each cortical canal being located near a furrow in the stem surface. This highly characteristic structure allows us to identify fossils related to Equisetum.

FIGURE 21-23 Sporangia of Equisetum occur in groups of 5 to 10 clustered together on a sporangiophore. Sporangiophores are grouped into strobili.

FIGURE 21-24 Calamites was an obvious relative to living equisetums, having jointed stems with whorls of leaves or branches at each node. A plant like this would have been a small tree about 10 m tall.
The evolutionary line of arthrophytes can be traced back through the trimerophytes to the rhyniophytes. In two groups of early arthrophytes, the Sphenophyllales and the Calamitales, a vascular cambium produced secondary xylem. Presumably secondary phloem was produced also, but the preservation of the fossils is not good enough to be certain (FIGURE 21-24). These plants became large trees up to 30 cm in diameter and more than 20 m tall. They had true monopodial growth, a main trunk, lateral branches, true leaves, and true roots.
The equisetophyte vascular cambium evolved independently from that of the lycophytes, yet the two suffered from the same defect: The fusiform initials could not undergo radial longitudinal division to produce more fusiform initials. As the wood accumulated and pushed the cambium outward, the arthrophyte fusiform initials finally became too large to function, and secondary growth ceased.
Although equisetophytes are an ancient clade with only a few surviving species, the plants are by no means primitive, poorly adapted, or rare. They occur worldwide in moist habitats and grow vigorously enough to be weeds in cultivated areas (FIGURE 21-25).

FIGURE 21-25 Although Equisetum lacks many derived features, it is by no means primitive or poorly adapted. Here it is a successful weed, competing well with both a corn crop and its human farmers.
Ferns
Aside from the equisetophyte line of evolution out of the trimerophytes, probably two more led to the plants that dominate Earth at present: ferns (TABLE 21-4) and woody plants (lignophytes, almost all of which are seed plants).
Early ferns first appeared in the Devonian Period and then diversified greatly (FIGURES 21-26 and 21-27). Whereas equisetophytes only have 15 species of living plants, the rest of the monilophytes have more than 12,000 species, and almost all of those are what are called leptosporangiate ferns (Leptosporangiatae; these are the ferns that are familiar to you). The other three groups of monilophytes (Marattiales, Ophioglossales, and Psilotales) have less than 300 species of living plants altogether, and fewer people will be familiar with them.
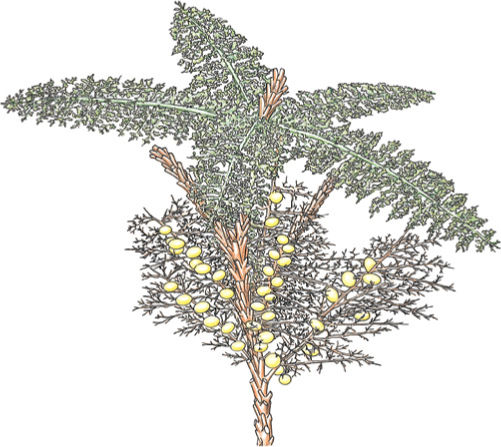
FIGURE 21-26 This reconstruction of a fern ancestor, Rhacophyton, shows spirally arranged branch systems that had evolved nearly to the point of being called leaves. Some are fertile with sporangia, and some are sterile; the former evolved into sporophylls, the latter into foliage leaves.
TABLE 21-4 Classification of Division Pteridophyta
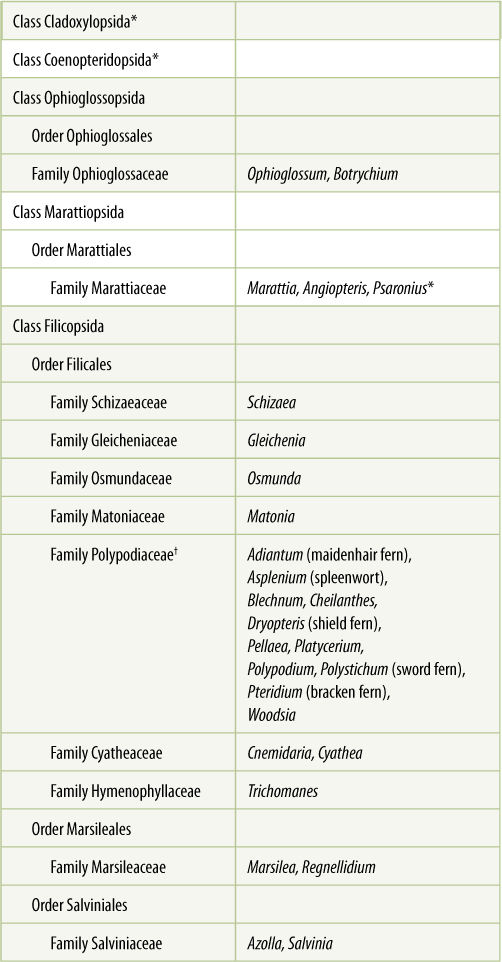
*All species in this group are extinct.
†This family contains almost all the living species of ferns.
Ferns can be found in almost any habitat. Moist, shady forests and lakesides are often considered “typical” fern habitats, but species of Woodsia and Cheilanthes occur in dry, hot deserts; Salvinia and Azolla grow floating on water; and Ceratopteris lives submerged below water (FIGURE 21-28). Other genera contain epiphytes (Polypodium) or vines (Lygodium). All ferns are perennial and herbaceous; none is woody, but some do achieve the size of small trees—the tree ferns Angiopteris, Cyathea, Cnemidaria, and others. Although called tree ferns, they never have secondary xylem.
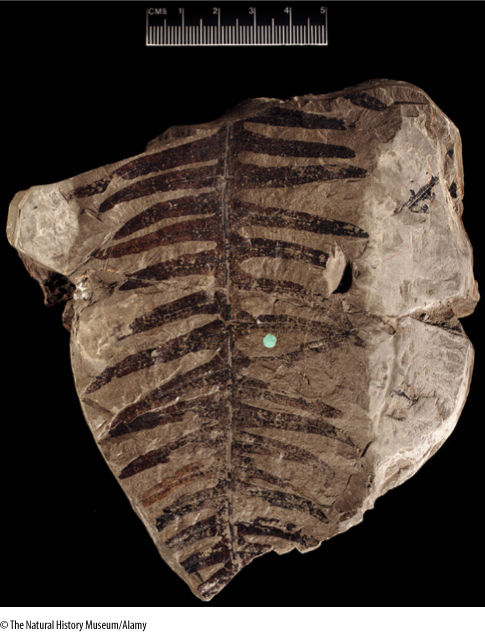
FIGURE 21-27 A later fern, Phlebopteris, from the Triassic Period. Even though the Triassic occurred from 230 to 190 million years ago, plants had evolved so many modern features that this fossil is not an ancestor to ferns in general, but to a particular family of living ferns, the Matoniaceae.
The fern sporophyte consists of a single axis, either a vertical shoot or a horizontal rhizome, that bears both true roots and megaphyllous leaves. The vascular system of the stem is an endarch siphonostele, a derived trait also present in equisetophytes and seed plants (FIGURE 21-29). At each node, a leaf trace diverges from the siphonostele, leaving a small segment of the vascular cylinder as just parenchyma; this region is a leaf gap. A vascular cambium has been reported to occur in one fern, Botrychium.

FIGURE 21-28 (A) Many ferns grow in hot, dry climates with little rainfall. If transplanted to habitats often described as typical for ferns—cool, moist, and shady—these would die within days. Azolla (B) and Salvinia (C) do not look like ferns at first glance, but the structure of their leaves and sporangia shows they are ferns. (D) Several genera of ferns such as this Cyathea are referred to as tree ferns because they have very long vertical trunks and huge fronds; however, unlike trees, these never have secondary growth and are actually giant herbs.
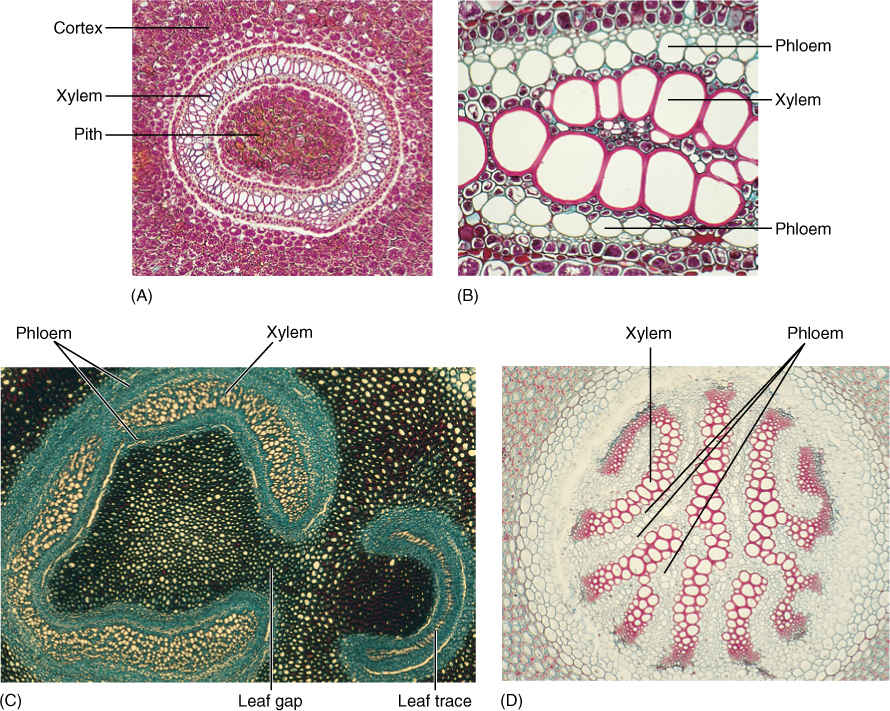
FIGURE 21-29 Anatomy of fern stems is variable—not only from species to species, but also within a single plant. (A) In this Dennstaedtia and many other ferns, xylem forms a complete ring in transverse section. (B) Phloem occurs on both the exterior and interior of the xylem in ferns; phloem interior to xylem is extremely rare in seed plants (× 20). (C) In this Adiantum, the vascular tissue makes almost a complete cylinder. A leaf gap occurs where vascular tissue moves to the leaves. Leaf gaps are characteristic of ferns; they do not occur in the microphyll line of evolution (lycophytes). (D) Members of the microphyll line of evolution, such as this Lycopodium annotinum, do not have pith (× 20).
Leaves of ferns may be leathery or delicate, only one cell thick in filmy ferns, but many layers thick in most. They have an upper layer of palisade parenchyma and a lower region of spongy mesophyll. Leaves are small (Trichomanes) or up to several meters long (tree ferns), but they are almost always compound with a rachis and leaflets (FIGURE 21-30). Staghorn fern (Platycerium) and bird’s-nest fern (Asplenium) have simple leaves. Fern leaf primordia have a distinct apical cell, unlike leaves of seed plants, and as the primordium grows, it curves inward, producing a tightly coiled young leaf commonly known as a fiddlehead (FIGURE 21-31). As the leaf expands, it uncoils and becomes flat. Fern leaves, especially larger ones, usually contain a considerable amount of vascular tissue. Leaf veins typically branch dichotomously.
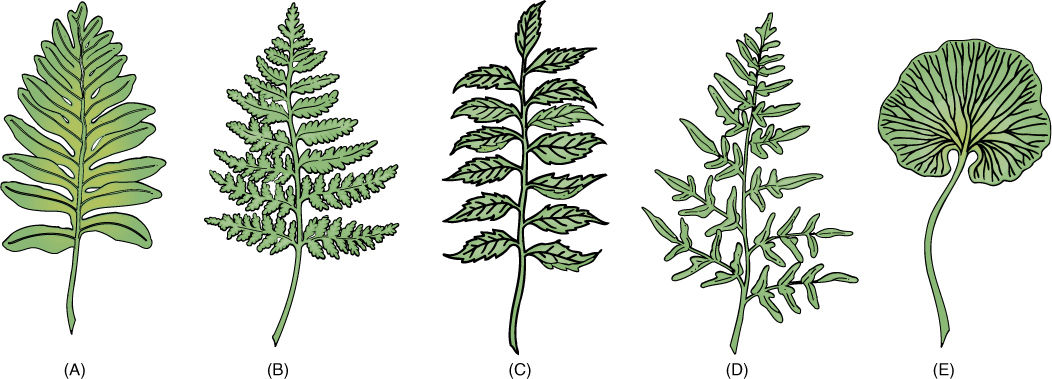
FIGURE 21-30 (A) — (E) Fern leaves (fronds) have a petiole and a lamina, the lamina being almost always pinnately compound. The midrib is a rachis, and the leaflets may also be compound.
Botany and Beyond
BOX 21-2 Form Genera
Although we can be fairly sure that a plant like Sciadophyton was the gametophyte of a plant like Rhynia or Cooksonia, we can probably never know for certain. These fossils are the separate pieces of a life cycle, just as the individual fossils of spores, leaves, fruits, flowers, stems, and roots are the pieces of plants. How do we know which fossil parts came from the same species? As a plant dies, its leaves and flowers usually abscise, and thus, we have numerous individual leaf and flower fossils mixed with the remains of bare stems. It is rare to find, at present or in the past, entire meadows or forests that consist of just a single species; usually many species grow mixed together. We find many types of fossil leaves mixed with many types of fossil stems, flowers, wood, and other plant parts. If we find a branch with leaves or flowers still attached, that is, if there is an organic connection, we can establish which organs are part of one plant.
Until we know which parts were portions of the same species, we must use form genera, which are created for types of isolated organs, tissues, spores, or pollen. If a fossil leaf is found which appears to be distinct from all others, it is named as a new form genus of leaf. As more fossil leaves are found in rocks of similar age, some appear to be related to the first and are assigned to that form genus. Mixed with the leaves may be twigs or large pieces of wood. We may be almost certain that the leaves have been produced by the twigs, based on this correlation of common occurrence at the same time and the same place; however, the leaves cannot be assigned to the form genus of the wood unless they are actually found still connected to a twig. Only when an organic connection has been established can two form genera be combined. For spores and pollen, no organic connection ever occurs, but occasionally, an unopened sporangium or anther is found; therefore, certain spores or pollen grains can be associated with those sporangia or anthers. Even when two form genera have been shown to be parts of the same species, it is still often much simpler to continue using the names of the form genera.
In some species, for example Blechnum spicant and cinnamon fern (Osmunda cinnamomea), certain leaves serve only for photosynthesis, and others serve only as sporophylls; however, in most ferns, each leaf does both (FIGURE 21-32). On the underside of the leaf are sori (singular, sorus), clusters of sporangia where meiosis occurs (FIGURE 21-33). Most ferns are homosporous; only two groups of water ferns (Marsileaceae and Salviniaceae) are heterosporous.

FIGURE 21-31 (A) Fern leaves (but not equisetophyte leaves) have a highly characteristic development: Young leaves are tightly coiled and uncurl as they expand. This is true not only of the rachis, but of the leaflets as well. This is circinate vernation. (B) While young like this, fiddleheads are edible and are cooked as a vegetable. Once mature, fern leaves are inedible.

FIGURE 21-32 (A) This cinnamon fern (Osmunda cinnamomea) contains both foliage leaves that never bear sporangia and highly distinct sporophylls that do bear sporangia. (B) Most ferns have only one type of leaf; the broad, thin shape of its leaflets is selectively advantageous for photosynthesis, and they bear clusters of sporangia (sori) on their underside. These sori are naked; they have no covering. (C) The sporangia of this maidenhair fern (Adiantum) occur near the margin of the leaflets and are protected by a flap of tissue called an indusium.
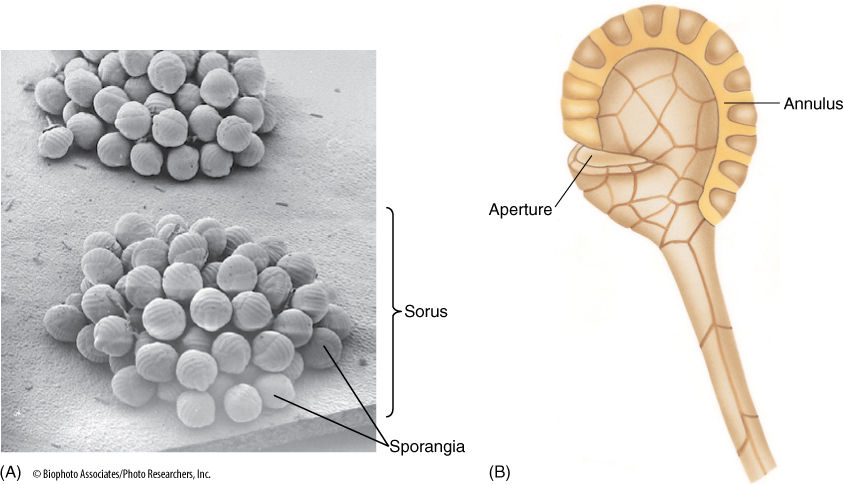
FIGURE 21-33 (A) A sorus contains many sporangia, each of which consists of a stalk and the actual sporangium body where meiosis occurs. (B) The sporangium opens when specialized cells of the annulus dehydrate, shrink, and suddenly crack open, expanding rapidly and throwing the spores (× 40).
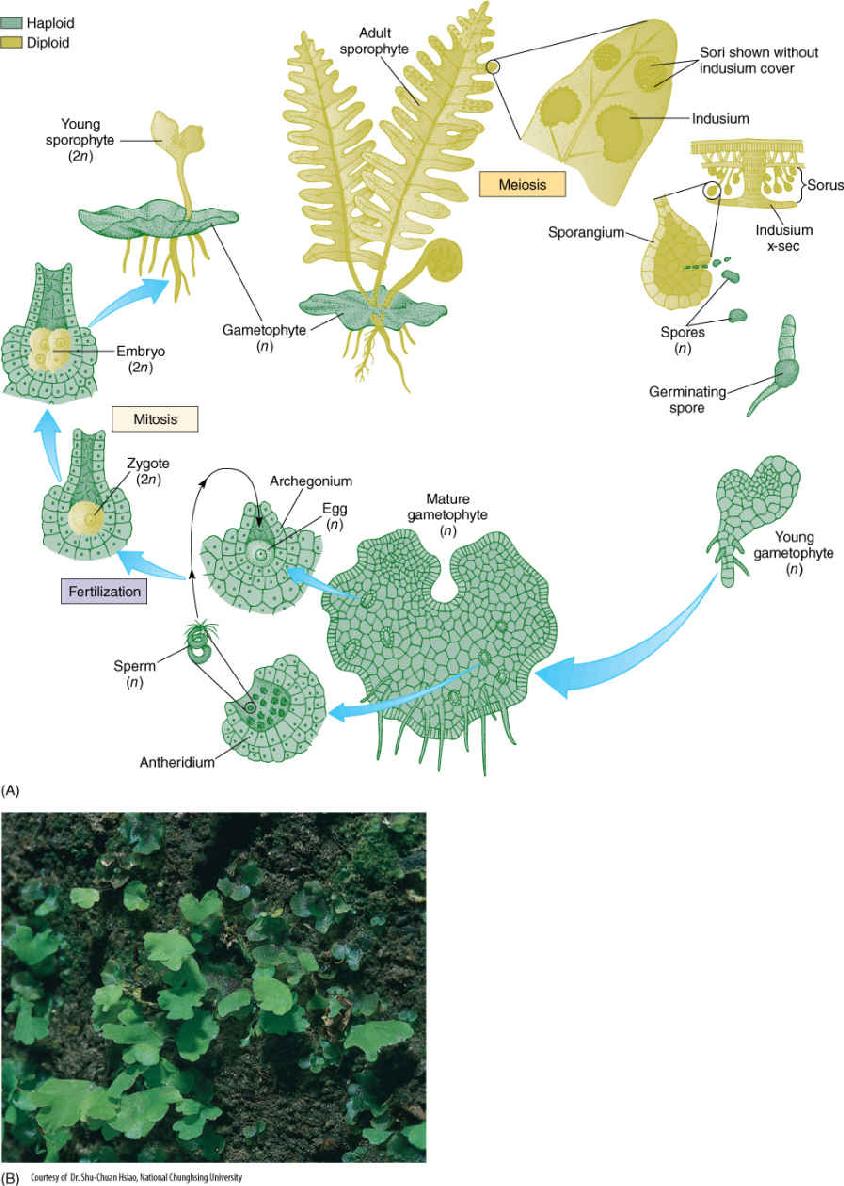
FIGURE 21-34 (A) The life cycle of a fern. Details are given in the text. (B) Fern gametophytes, several with the first leaves of young sporophytes.
When fern spores germinate, they grow into small, simple heart-shaped or ribbon-shaped photosynthetic gametophytes with unicellular rhizoids on the lower surface but with no vascular tissue and no epidermis (FIGURE 21-34). Each usually bears both antheridia and archegonia. Antheridia develop early and can be difficult to detect because they occur among rhizoids; archegonia develop later, close to the gametophyte apex. When the environment is sufficiently moist, antheridia release motile sperm cells that could easily be mistaken for unicellular green algae except for the absence of plastids. These swim to the archegonia and fertilize the egg. As in Equisetum, a fern zygote is retained by the gametophyte that nourishes it; however, the young sporophyte soon produces its first leaves and roots and becomes independent. Its continued growth eventually destroys the small gametophyte.
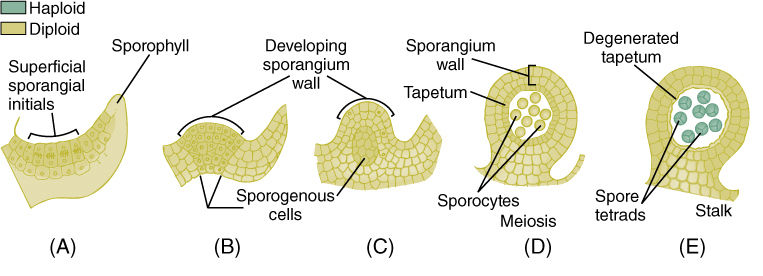
FIGURE 21-35 Development of eusporangia. (A) Surface cells undergo divisions and the inner daughter cells become sporogenous. (B) — (E) Further divisions result in a large number of spores and a thick sporangium wall.
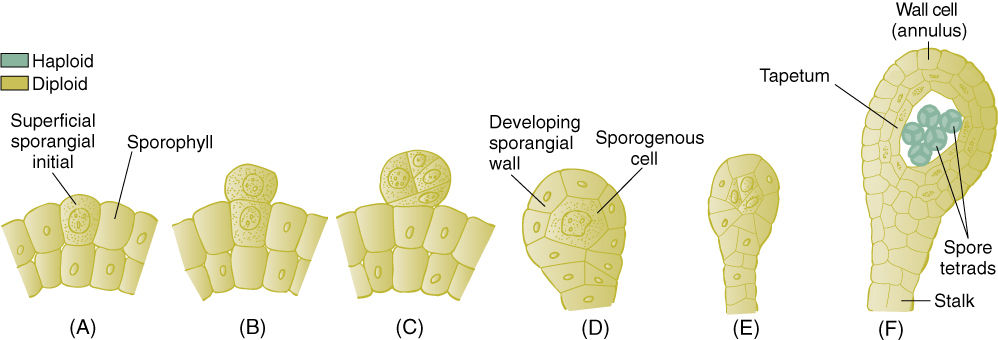
FIGURE 21-36 Development of leptosporangia. (A) and (B) The sporangium is initiated by division in just a single surface cell. (C) — (F) After a few cell divisions, the sporangium has a stalk, a thin wall, and only a few spores.
Eusporangia and Leptosporangia
Ferns contain two types of sporangia that differ in fundamental aspects of their development. The eusporangium is initiated when several surface cells undergo periclinal divisions, resulting in a small multilayered plate of cells (FIGURE 21-35). The outer cells develop into the sporangium wall, and the inner cells proliferate into sporogenous tissue. This results in a relatively large sporangium with many spores. This is the fundamental type of sporangium, and it exists in virtually all plants other than Leptosporangiate; even the anthers and ovules of flowering plants develop this way.
Leptosporangia are initiated when a single surface cell divides periclinally and forms a small outward protrusion (FIGURE 21-36). This undergoes several more divisions, which result in a small set of sporogenous cells and a thin covering of sterile cells; only a few spores are produced. Having leptosporangia is a derived trait occurring only in the clade Leptosporangiatae.
Psilotum and Tmesipteris
These two small genera contain the simplest of all living vascular plants (FIGURE 21-37). Psilotum is constructed very much like Rhynia and Aglaophyton, and until the mid 1970s, they were often placed together, even though Rhynia and Aglaophyton are extinct. Plants of Psilotum (“whisk ferns”) are now known to be highly derived, highly simplified ferns. They are small plants with prostrate rhizomes and upright stems that branch dichotomously; they have an epidermis, cortex, and a simple vascular cylinder with no pith—a protostele (FIGURE 21-38). Xylem consists of annularly or helically thickened tracheids. Psilotum is unique among living vascular plants in that it has lost the capacity to make roots and leaves. Shoots do have occasional small, scale-like projections of tissues resembling enations.

FIGURE 21-37 The growth habit of Psilotum stems—dichotomous axes with eusporangia.
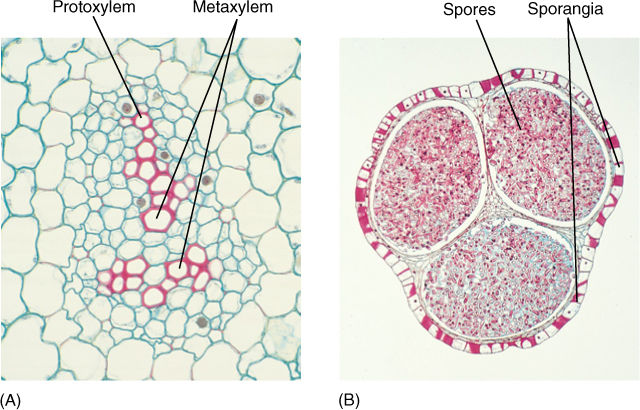
FIGURE 21-38 (A) Vascular structure of Psilotum is simple: a protostele and phloem surrounding the xylem. Protoxylem is exarch (to the exterior of the metaxylem), which is similar to the zosterophyllophytes rather than the rhyniophytes (×100). (B) Sporangia of Psilotum are actually considered to be three sporangia fused together; each is at the end of an extremely short branch, basically a three-branched shoot subtended by a scale (× 8).
Particularly unusual are the gametophytes, short, branched cylinders less than 2 mm in diameter. Their surface is covered with elongate cells, rhizoids, that anchor and perhaps also absorb (FIGURE 21-39). Gametophytes have no chlorophyll but instead are heterotrophic, forming either a symbiotic or a parasitic association with soil fungi; fungi invade most cells of the gametophyte and provide it with sugars and minerals. Psilotum is the only species whose gametophytes contain vascular tissue: There is a small amount of vascular tissue similar to that of sporophytes. A central mass of tracheids is surrounded by phloem and an endodermis.
Psilotum occurs in tropical and subtropical regions. In the United States, it can be found in the Gulf Coast states from Florida to Texas, as well as in Hawaii. Tmesipteris is limited to Australasia, primarily Australia and other South Pacific islands.
![]() The Term “Vascular Cryptogams”
The Term “Vascular Cryptogams”
Traditionally, the plants of this chapter have been referred to informally as vascular cryptogams (or often as “ferns and fern allies”). This name indicates that they have vascular tissue and that because they lack seeds their reproduction is hidden (crypto). Although these are two important features, they are shared ancestral features—symplesiomorphies—and thus do not indicate they are closely related. What is most striking about these plants are the many features they lack: seeds, flowers, fruits, and so on; however, we construct cladograms on the basis of shared derived features—synapomorphies—not on a lack of features, and consequently, we do not group vascular cryptogams together formally. Also, if we tried to consider vascular cryptogams as a clade made up of several subclades (lycophytes and monilophytes for example), we would be creating a group that contains an ancestral group but not all its descendants: The ancestor of the monilophyte line is also the ancestor of lignophytes. To leave lignophytes out would be to create a paraphyletic group, which is not correct. Instead, when we use a term such as “vascular cryptogam,” we are referring to a grade, to basically a level of evolutionary advancement. Similarly, the term “bryophyte” is often used for all the nonvascular plants, not to indicate that they are related, but that they are at a level of evolution that does not include the presence of vascular tissue.
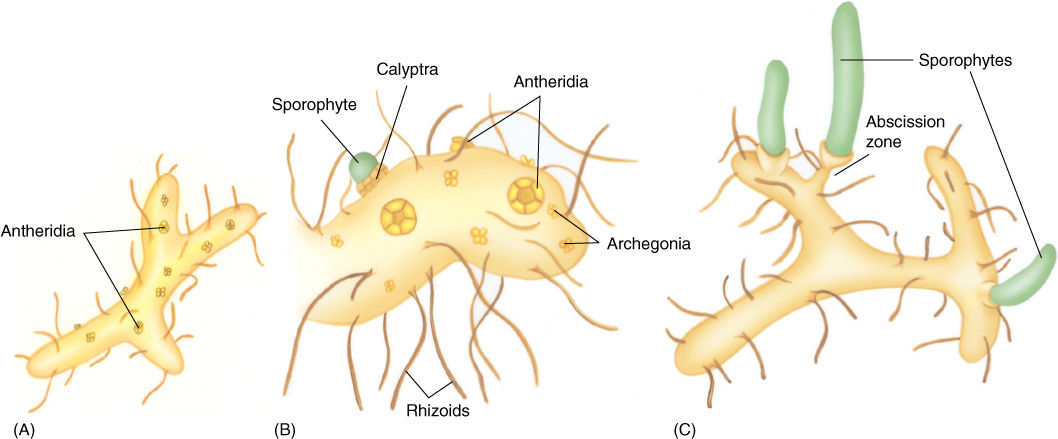
FIGURE 21-39 Gametophytes of Psilotum are small cylinders (A) that live heterotrophically, nourished by endophytic soil fungi. (B) Each gametophyte bears both antheridia and archegonia, and because the egg and zygote are retained in the archegonium, the young sporophyte initially is nourished by the parent gametophyte; during early development, there is proliferation of gametophyte tissue as a protective calyptra. (C) As the sporophyte grows and enlarges, it forms an abscission zone and detaches from the gametophyte, becoming independent.
 At the Next Level
At the Next Level
1. Zosterophyllophytes and lycophytes. It is difficult to imagine but the plants that surround us now are in some respects the second flora of Earth. After true plants diverged from algae, the zosterophyllophytes and their descendants the lycophytes dominated Earth and formed extensive forests. This chapter gives only the basics of their biology, and most of what we know about them is their anatomy, some details of their reproduction, and almost nothing of their physiology (Did they have C4 photosynthesis? Associations with nitrogen-fixing bacteria?). Unfortunately, there is no book on zosterophyllophyte biology, but there are review papers and summaries. If you choose these plants for further study, be certain to look up information about the climate at that time: The atmosphere had much more carbon dioxide and it was much warmer than now.
2. The fern life cycle. All plants have a life cycle in which diploid sporophytes alternate with haploid gametophytes. But in most plant groups, this just seems to be something to be memorized, something abstract: It never seems real. But with ferns, we can easily cultivate both generations of the life cycle and have an experience in which we truly understand it. For this independent study, find a fern with sporangia on the underside of its leaves. Collect spores by simply laying the leaf on a sheet of paper overnight. If there are no spores on the paper after several hours, the leaf might be too old and you will need to use a younger one. Sprinkle the spores onto moist potting soil in a plastic flower pot. Cover with plastic wrap then place the pot in bright, indirect light. Keep it moist by placing it in a deep dish of tap water once a week or so. After 2 or 3 weeks, the potting soil should turn green with gametophytes—just threadlike at first, then heart-shaped after another 2 or 3 weeks. They often grow so thickly that the entire surface of the potting soil becomes covered with gametophytes. After they have grown to the heart-shaped stage, take off the plastic wrap, pour water on to the gametophytes, then cover again and wait. You have just helped them have sex, and sperm cells should be swimming from antheridia to archegonia; in a week or two, the embryos will produce their first leaves. After another month or so you can transplant some sporophytes into individual flower pots. You will have encountered all stages the plant life cycle firsthand.
SUMMARY
1. Some early gametophytes were the most elaborate ever and subsequent gametophyte evolution from them has involved simplification and reduction.
2. Lycophytes and monilophytes (vascular cryptogams) are embryophytes, not algae, because of their multicellular, jacketed sporangia, and gametangia. They have vascular tissue, and thus, they are not mosses, liverworts, or hornworts. Although some have small endosporial megagametophytes, none is retained in the sporangium; therefore, none is a seed.
3. The earliest vascular plants are rhyniophytes, plants with simple dichotomously branched stems, no leaves, and endarch protoxylem.
4. Zosterophyllophytes were similar to rhyniophytes but had exarch protoxylem and lateral sporangia that opened across their tops. Zosterophyllophytes probably were the ancestors to lycophytes.
5. Three types of leaf occur in the plant kingdom: gametophyte leaves of nonvascular plants, microphylls of lycophytes, and megaphylls of all nonlycophyte vascular plants. Microphylls evolved from enations; megaphylls are modified telomes—branch systems.
6. Vascular cambia evolved at least three times; cells in the two that evolved in ancient lycophytes and equisetophytes could not divide with a radial longitudinal wall.
7. Trimerophytes were transitional between early vascular plants and euphyllophytes. Evolutionary advances in the trimerophytes were pseudomonopodial branching and the first steps in megaphyll evolution.
8. Equisetophytes contain only one small genus of living, herbaceous species and several extinct genera of large, complex, tree-like plants. All had a characteristic jointed structure and various systems of canals.
9. Ferns have many living species and tremendous diversity. No ferns have secondary growth or seeds, but all have megaphylls.
IMPORTANT TERMS
cones
enations
endarch protostele
equal dichotomous branching
euphyllophytes
exarch protostele
horsetails
leaf gap
leaf trace
lignophytes
megaphylls
microphylls
monilophytes
monopodial growth
overtopping
planation
protostele
rhyniophytes
scouring rushes
seed plants
siphonostele
sorus
sporophyll
strobili
telome theory
vascular cryptogams
webbing
zosterophyllophytes
REVIEW QUESTIONS
1. The earliest fossils that definitely were vascular land plants belong to _________________, a genus of extinct plants. They branched _______________, both branches being of equal size and vigor.
2. Draw a plant of Rhynia. Be certain to include the reproductive parts. Now assume that it has a life cycle with an alternation of isomorphic generations and draw a complete life cycle.
3. If Rhynia or its contemporaries were the ancestors to the ferns, how did the gametophytes and sporophytes change during evolution?
4. In the vascular bundles of flowering plants, protoxylem is closest to the center of the stem, and metaxylem is farther out. Is this an endarch or an exarch arrangement? Seed plants always have just this one arrangement, but what about the early vascular plants. Were they endarch or exarch, or did both types occur originally?
5. The vascular bundles of flowering plants surround a pith, but the earliest vascular plants had no pith. A vascular system with a solid mass of xylem with no pith is called a _________________. A stele that does have a pith is called a _________________.
6. Rocks that contain fossils of Rhynia also often contain fossils of Sciadophyton. We know that Rhynia was a sporophyte and that Sciadophyton was a gametophyte. Because they grew together, we suspect that they may be the two generations of a single species. If so, did Rhynia have an alternation of isomorphic or heteromorphic generations? Also, if this is true, evolution has caused one generation to become more complex, the other to become less complex. Which is which? Explain.
7. There are two alternative hypotheses about the life cycle of the early vascular plants, the transformation hypothesis and the interpolation hypothesis. If Rhynia and Sciadophyton are the two generations of one species, which hypothesis would be favored? Briefly describe the other hypothesis.
8. What are the zosterophyllophytes? How did they differ from rhyniophytes? Why do we think they are related to lycophytes but not to ferns and seed plants?
9. The reconstruction of Asteroxylon in Figure 21-10 shows thin, leaf-like flaps of tissue on the plant’s surface. What are these called? Did they ever have stomata in any of the zosterophyllophytes? Did they have vascular bundles? Did they ever become large (Hint: Look at Figure 21-13)?
10. What are microphylls? Are they related to the enations of Asteroxylon?
11. The lycophytes once contained many species of large trees that formed extensive forests. Briefly describe plants of Lepidodendron and Sigillaria. Also describe their wood.
12. Name two genera of living lycophytes. What are their common names? About how big do they get? Are they leafy or do they have naked stems? In a plant identification book, they would probably be listed with ferns.
13. Trimerophytes were plants that probably evolved from rhyniophytes but with more derived features. In one feature, certain stems grow longer than others, and thus, rather than having dichotomous branching, they have ____________ branching (displayed especially by Pertica). Simultaneously, the positioning of branches became more ______________ and _______________.
14. Describe the evolution of megaphylls. What are telomes?
15. Describe the trimerophytes. From what group did they evolve, and what lines of evolution did they produce? Even though all rhyniophytes and trimerophytes are now extinct, would you consider them unsuccessful?
16. What are the two common names of equisetums? What is the appearance of the plants? Their approximate height? Look at Figures 21-21 and 21-24. Equisetums have strobili (in plant identification books, these will be called cones) and canals. Briefly describe the canals and strobili.
17. Ferns first appeared in the Devonian Period. Look at the inside of the back cover. How long ago was the Devonian Period? Unlike all the groups mentioned so far in this chapter, most of this group is still extant, not extinct. About how many species of modern (i.e., not extinct) ferns are there?
18. Name two genera of ferns that are found in deserts. Name two that grow floating on water. One that lives underwater. There are ferns called “tree ferns.” Do tree ferns have woody trunks with secondary xylem?
19. A very important feature of ferns is shown in Figure 21-29: Their shoot xylem is not solid as it was in the rhyniophytes. In the evolution of ferns and seed plants, there was the evolution of pith. A stele with a pith is called a _________________. Another significant feature of ferns is that the phloem occurs both to the outer side as well as the inner side of the xylem. Where does it occur in the vascular bundles of flowering plants?
20. You may have noticed that the undersides (but never the upper sides) of fern leaves have brown dots or brown streaks or brown patches. The brown dots are called _________________ (singular, _________________), clusters of _________________ where meiosis occurs. If a leaf carries these structures, the leaf is called a ___________________.
21. When fern spores germinate, they grow into small, simple heart-shaped or ribbon-shaped ______________ with unicellular _________________ on the lower surface but with no vascular tissue and no _________________.
22. Briefly describe eusporangia and leptosporangia. Which is the type that occurs in all other vascular plants? Which is the type that occurs in most ferns?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green Plant Cells: © ShutterStock, Inc./Nataliya Hora; Purple Tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./Danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant Icon: © ShutterStock, Inc./Vector; Digging Man Icon: © ShutterStock, Inc./Z-Art