Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Plant Structure
Tissues and the Primary Growth of Stems

Chapter Opener Image: These strange little plants are seedlings of a desert-adapted species, Hoodia gordonii. Even at this young stage each has most of the basic plant tissues that protect the body, conduct materials through it, carry out photosynthesis, and so on. At the appropriate time, each will also make wood and bark, and also flowers, fruits, and seeds. These seedlings are being cultivated in a greenhouse, but the species is native to deserts in southwest Africa.
OUTLINE
✵ Concepts
✵ Basic Types of Cells and Tissues
- Parenchyma
- Collenchyma
- Sclerenchyma
✵ External Organization of Stems
✵ Internal Organization of Stems: Arrangement of Primary Tissues
- Epidermis
- Cortex
- Vascular Tissues
- Xylem
- Phloem
- Vascular Bundles
✵ Stem Growth and Differentiation
Box 5-1 Plants and People: Parenchyma, Sclerenchyma, and Food
Box 5-2 Plants Do Things Differently: Organs: Replace Them or Reuse Them?
Box 5-3 Alternatives: Simple Plants
Box 5-4 Plants Do Things Differently: Plants and People Grow Differently
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Summarize the evolution of land plants.
✵ Give examples of herbs and woody plants.
✵ Define the three classes of plant cells.
✵ Describe the components of basic stem organization.
✵ Provide examples of modifications to basic stem components.
✵ Recall the components of the internal organization of stems.
✵ State the functions of the epidermis.
✵ Contrast the way water moves through tracheids as compared to vessel elements.
✵ Order the processes of primary growth.
 Did You Know?
Did You Know?
✵ All soft parts of plants, such as flowers and leaves, are composed of cells that live only briefly, just a few days for flowers, just from spring until autumn for most leaves.
✵ Most hard parts of plants, such as wood and the shells around seeds, are composed of cells that usually die immediately after they have hardened.
✵ Almost all shoots, even the highly modified shoots of cacti, orchids, bulbs, and vines, have just one pattern of tissues: Once you have learned the pattern of one stem, you know the pattern present in almost all.
![]() Concepts
Concepts
The body of an herb contains just three basic parts: leaves, stems, and roots (FIGURE 5-1). When the first land plants evolved about 420 million years ago, they were basically just algae that either washed up onto a shore or were left there as lakes and streams evaporated. They had no roots, stems, or leaves, and they just lay on the mud. As the shores gradually became crowded with such plants, some grew over others, shading them. Any plant that had a mutation allowing it to grow upright into the sunshine above the others had a selective advantage. However, being upright is not easy: Elevated cells are out of contact with the moisture of the mud; therefore, water must be transported up to them. Elevated tissues act as a sail and tend to blow down, so supportive tissue is necessary. Absorptive cells in mud are shaded and cannot photosynthesize; thus, sugars must be transported down to them. Shortly after plants began living on land, distinct, specialized tissues and organs began to evolve.
As early populations of land plants continued to evolve and became taller, their stems functioned primarily as transport and support structures, as they do still. But, stems of modern flowering plants have additional roles. They produce leaves and hold them in the sunlight, and during winter, they store sugars and other nutrients, such as the sugary sap of maples. Stems may also be a means of survival: Underground bulbs and corms remain alive when above-ground leaves die. Stems of many species are a means of dispersal. They spread as runners or vines, or pieces break off and are carried by animals or water to new areas where they sprout roots and grow into new plants.
Although all flowering plants possess leaves, stems, and roots, these parts have been modified so extensively in some species that they may not be recognizable without careful study. For example, cacti are often described as leafless, but they actually have small green leaves between 100 and 1000 mm long (FIGURE 5-2). Large, broad leaves would be selectively disadvantageous for these desert plants because the extensive surface area of such leaves gives up so much water to the dry air that the plant would desiccate.
Similarly, all flowering plants have stems, but in some, they are only temporary, reduced structures. Orchids such as Campylocentrum pachyrrhizum and Harrisella porrecta consist of a mass of green photosynthetic roots connected to a tiny portion of stem; roots constitute almost the entire plant body (FIGURE 5-3). The shoot becomes active only when flowers are to be produced. One hypothesis is that this unusual body evolved because the ancestors of these species had roots that were more resistant to water stress than their stems were. Such plants could occupy drier habitats if either of two things happened: (1) Mutations were selected that caused stems and leaves to be more water conserving—this happened in most orchid species (FIGURE 5-4), or (2) mutations occurred that enhanced the root’s ability to absorb carbon dioxide and carry out photosynthesis. Although thousands of plant species can withstand harsh conditions, only a handful do so by being “shootless” and having photosynthetic roots. Mutations that permit this type of body may be either rare or generally detrimental.
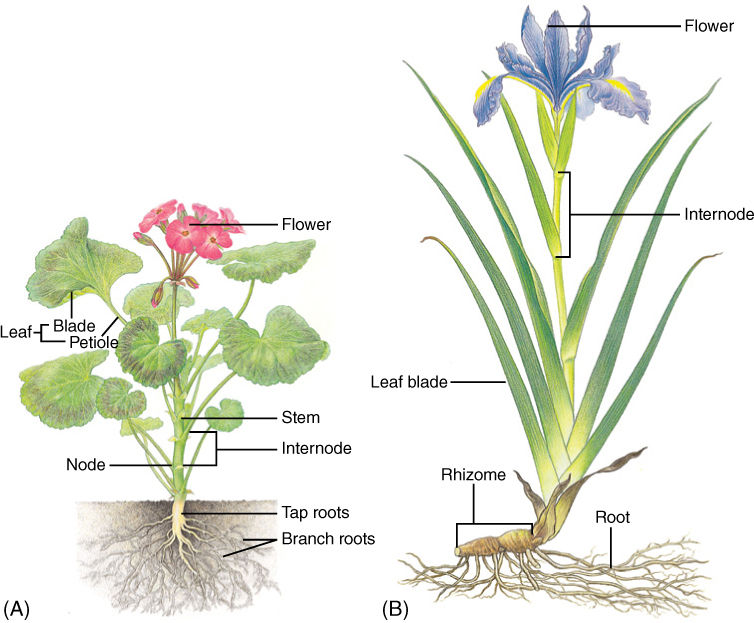
FIGURE 5-1 (A) The primary body of an herb like this geranium consists of roots, stems, and leaves; buds are located in the axil of each leaf and may grow to be either a vegetative branch or a set of flowers. (B) This Iris is also an herb and never produces wood or bark. Almost all flowering plants are either broadleaf plants (eudicots) like the geranium or plants with grass-like leaves (monocots) like the Iris.

FIGURE 5-2 This prickly pear (Opuntia) shows that one plant can have two types of shoot: The “pad” is the main shoot, and the spine clusters are highly modified axillary branch shoots. One of the spine-bearing branches has been stimulated to develop into the first type of shoot and become a pad-like branch. The plant also has two types of leaves: (1) small fleshy green leaves on the young buds and (2) spines on the axillary branches.
Some plants in the bromeliad family are nearly rootless. In the coastal deserts near Lima, Peru, fog is frequent, but rain never falls. Because the soil is always dry, roots are of little use. Plants of Tillandsia straminea of that region are small, herbaceous vines that lie on top of the soil (FIGURE 5-5). Plants absorb moisture through leaves made wet by fog, and they derive minerals from wind-blown dust that dissolves on moist leaf surfaces. The plants are not anchored to the soil but roll over coastal dunes as the wind blows them. Such a lifestyle would be impossible for a tree or bush because large plants could not absorb enough water or minerals without roots.

FIGURE 5-3 This orchid carries out photosynthesis by means of roots that contain chlorophyll. The plant grows in trees, attached to their bark (it is epiphytic) such that its roots are exposed to sunlight, they are never in soil. The photosynthetic roots grow in any direction, even upward. The plant has only a small portion of shoot, which produces flowers.

FIGURE 5-4 Most epiphytic orchids resist the stresses of temporary drying because their shoots are fibrous and have a thick cuticle composed of cutin and wax.
In each plant described, leaves, stems, or roots have become highly modified by natural selection, permitting survival in unusual habitats; however, in no species have any of these organs been completely lost evolutionarily. We must assume that the organ carries out some essential function. In “shootless” orchids, the residual shoot is necessary for flowering and sexual reproduction. Cactus leaves are involved in the formation of buds that produce the defensive spines. Although roots of T. straminea neither absorb water and minerals nor anchor the plant, they may be essential for production of critical hormones, as are roots of other plants. In all cases, analysis of many aspects of the plant’s biology reveals how the structure and metabolism of a particular organ are adaptive, how modifications affect other plant parts, and how organs have subtle functions not always obvious in more “typical” plants.

FIGURE 5-5 These bromeliads are not rooted into the coastal sand dunes—a strong wind can blow them around. All water is absorbed from fog condensing on the leaves.
TABLE 5-1 Types of Plant Body
The flowering plants discussed here are formally classified as the division Magnoliophyta, but they are known informally as angiosperms. This group consists of approximately 297,000 species and is the largest division in the plant kingdom. Early angiosperms diversified into several groups that are now known as basal angiosperms, eudicots, and monocots. Most basal angiosperms probably will not be familiar to you except for waterlilies, magnolias, and laurels. Eudicots are broadleaf plants such as roses, asters, maples, and others, and monocots are grasses, lilies, cattails, palms, philodendrons, bromeliads, and several others.
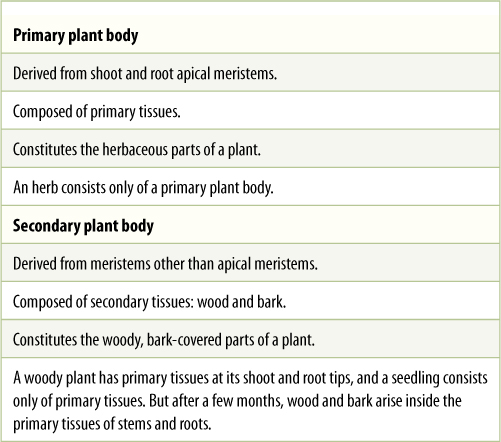
This chapter began by stating that the body of an herb contains three parts; it is necessary to explain more precisely what an herb is. Plant bodies are of two fundamental types: an herbaceous body, also called a primary plant body, and a woody body, known as a secondary plant body. An herb is a plant that never becomes woody and covered with bark; it often lives for less than a single year (snapdragons, petunias, beans, corn, and wheat), although many live and grow for years (irises and lilies). Its tissues are primary tissues. In woody plants such as trees and shrubs (oaks, maples, magnolias, roses, and boxwood), the wood and bark are secondary tissues (TABLE 5-1). A few plants are surprising: Palm trees are large, perennial, and very hard, but they do not actually have wood; thus, they are giant herbs, not woody plants.
![]() Basic Types of Cells and Tissues
Basic Types of Cells and Tissues
Despite the diversity of types of stems that have originated by natural selection, all share a basic, rather simple organization. The same is true for leaves and roots. Although we might suspect that a plant contains numerous types of cells, actually, all plant cells are customarily grouped into just three classes based on the nature of their walls: parenchyma, collenchyma, and sclerenchyma.
Parenchyma
Parenchyma cells have only primary walls that remain thin (TABLE 5-2). Parenchyma tissue is a mass of parenchyma cells. This is the most common type of cell and tissue, constituting all soft parts of a plant. Soft leaves, petals, fruits, and seeds are composed almost completely of parenchyma. Parenchyma cells are active metabolically and usually remain alive after they mature. Numerous subtypes are specialized for particular tasks (FIGURE 5-6).
Chlorenchyma cells are parenchyma cells involved in photosynthesis; they have numerous chloroplasts, and their thin walls allow light and carbon dioxide to pass through to the chloroplasts. Other types of pigmented cells, as in flower petals and fruits, also must be parenchyma cells with thin walls that permit the pigments in the protoplasm to be seen.
Glandular cells that secrete nectar, fragrances, mucilage, resins, and oils are also parenchyma cells; they typically contain few chloroplasts but have elevated amounts of dictyosomes and endoplasmic reticulum. They transport large quantities of sugar and minerals into themselves, transform them metabolically, and then transport the product out.
Transfer cells are parenchyma cells that mediate shortdistance transport of material by means of a large, extensive plasma membrane capable of holding numerous molecular pumps. Unlike animal cells, plant cells cannot form folds or projections of their plasma membranes; instead, transfer cells increase their surface area by having extensive knobs, ridges, and other ingrowths on the inner surface of their walls (FIGURE 5-7). Because the plasma membrane follows the contour of all these, it is extensive and capable of large-scale molecular pumping.
Some parenchyma cells function by dying at maturity. Structures such as stamens and some fruits (such as pea pods) must open and release pollen or seeds; the opening may be formed by parenchyma cells that die and break down or are torn apart. Large spaces are often needed inside the plant body, for example, to allow gases to diffuse within leaves (Figure 5-6B and Figure 5-10E); many such spaces are formed when the middle lamella decomposes, and cells are released from their neighbors. In other cases, the space is formed by the degeneration of parenchyma cells. In a few species, such as milkweeds, as parenchyma cells die, their protoplasm is converted metabolically into mucilage or a milky latex.
TABLE 5-2 Three Basic Types of Plant Cells and Tissues, Based on Cell Wall

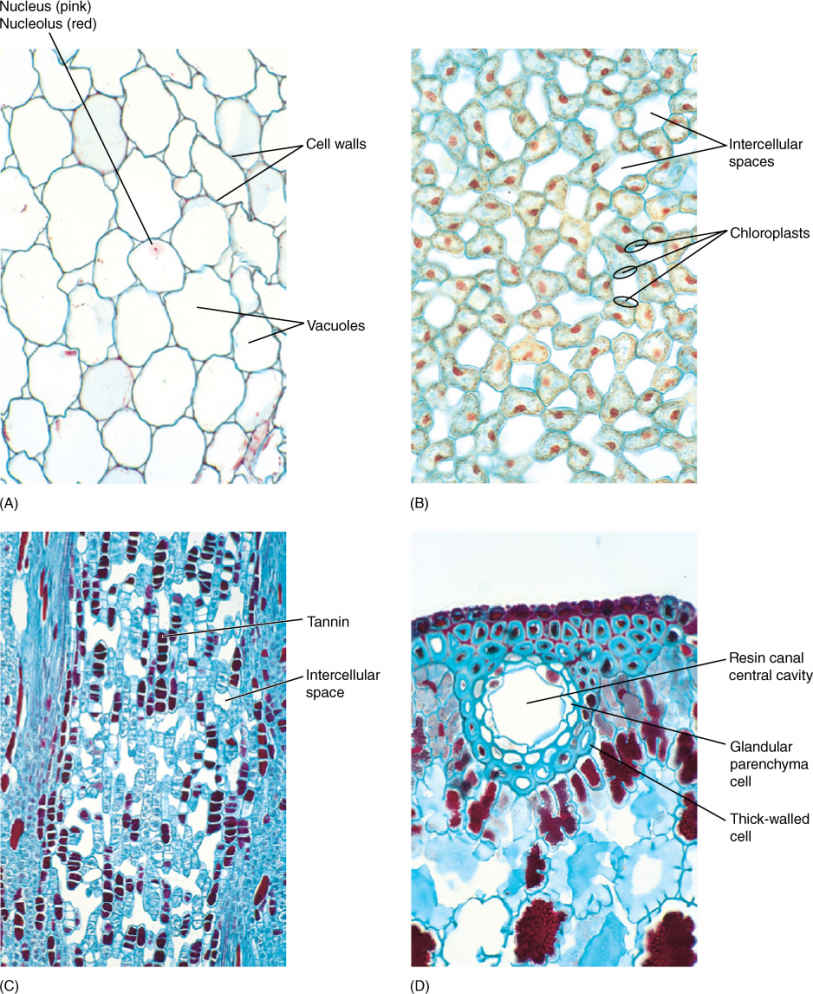
FIGURE 5-6 (A) Parenchyma cells of geranium. Their walls (blue green) are thin, and their vacuoles are large and full of watery contents that did not stain. Nuclei were present in all cells, but because these cells were so large and the section (slice) was cut so thin, most nuclei were cut away during the preparation of this slide. One nucleus is still present (×160). (B) Chlorenchyma cells from a leaf of privet. Because these cells are small and the section is thicker than that in (A), most cells still have nuclei (red). The structures close to the wall (blue) are chloroplasts. The large white areas are intracellular spaces where the cells have pulled away from each other, permitting carbon dioxide to diffuse rapidly throughout the leaf (×160). (C) Material taken from the center of a pine (Pinus) stem. Cells that have stained dark purple are filled with chemicals called tannins; these are bitter and deter insects from eating the tissue (×50). (D) A resin canal in a pine leaf. The white area is the central cavity where resin is stored, and cells that line the cavity are glandular parenchyma cells that synthesize and secrete resin. The innermost cells have thin walls, which permit movement of resin from the cells to the cavity. The outer cells have thick walls, which provide strength; cells with thick walls are not parenchyma cells (×160).
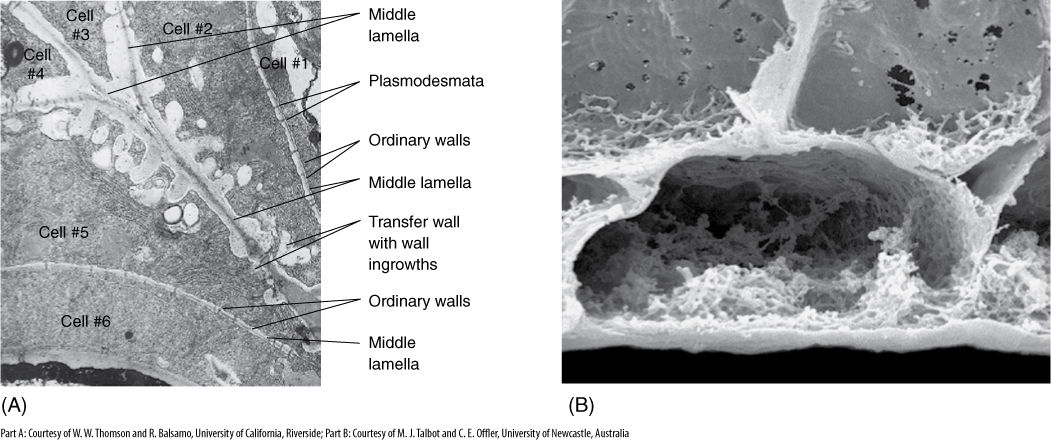
FIGURE 5-7 (A) Transfer cells in the salt gland of Frankenia grandifolia. The wall ingrowths increase the surface area of the cell membrane, providing more room for salt-pumping proteins in the membrane (×20,000). (B) These transfer cells were frozen, then broken open, and all of the protoplasm was washed away with detergent. Only the cell wall is left. All the knobby irregular areas are wall ingrowths (×5,000).
Parenchyma tissue that conducts nutrients over long distances is phloem; it is discussed later in this chapter.
Parenchyma cells are relatively inexpensive to build because little glucose is expended in constructing the cellulose and hemicellulose of such thin walls. Most parenchyma cell walls are only 80 to 100 nm thick, with just 5 to 10 layers of cellulose microfibrils. Each molecule incorporated into a wall polymer cannot be used for other functions such as generating ATP or synthesizing proteins. Consequently, it is disadvantageous to use a cell with thick walls any time one with thin walls would be just as functional. Most leaves are soft, composed almost entirely of parenchyma, and are therefore not very expensive metabolically. After several weeks of photosynthesis, they replace the sugar used in their construction, and all photosynthesis after that point is net gain for the plant.
Collenchyma
Collenchyma cells have a primary wall that remains thin in some areas but becomes thickened in other areas, most often in the corners (FIGURE 5-8). The nature of this wall is important in understanding why it exists and how it functions in the plant. Like clay, the wall of collenchyma exhibits plasticity, the ability to be deformed by pressure or tension and to retain the new shape even if the pressure or tension ceases. Collenchyma is present in elongating shoot tips that must be long and flexible, such as those of vining plants like grapes. It is present as a layer just under the epidermis or as bands located next to vascular bundles, making the tips stronger and more resistant to breaking (FIGURE 5-9); however, the tips can still elongate because collenchyma can be stretched. In species whose shoot tips are composed only of weak parenchyma, the tips are flexible and delicate and often can be damaged by wind; the elongating portion must be very short, or it simply buckles under its own weight.
It is important to think about the method by which collenchyma provides support. If a vine or other collenchyma-rich tissue is cut off from its water supply, it wilts and droops; the collenchyma is unable to hold up the stem. Parenchyma cells are needed in the inner tissues for support. Collenchyma and turgid parenchyma work together like air pressure and a tire: The tire or inner tube is extremely strong but is useless for support without air pressure. Similarly, air pressure is useless unless it is confined by a container. In stems, the tendency for parenchyma to expand is counterbalanced by the resistance of the collenchyma, and the stem becomes rigid but able to grow.
Because collenchyma cell walls are thick, they require more glucose for their production. Collenchyma is usually produced only in shoot tips and young petioles, where the need for extra strength justifies the metabolic cost. Subterranean shoots and roots do not need collenchyma because soil provides support, but aerial roots of epiphytes such as orchids and philodendrons have a thick layer of collenchyma.
Sclerenchyma
The third basic type of cell and tissue, sclerenchyma, has both a primary wall and a thick secondary wall that is almost always lignified (FIGURE 5-10). These walls are elastic: They can be deformed, but they return to their original size and shape when the pressure or tension is released. Sclerenchyma cells develop from parenchyma cells in mature organs after they have stopped growing and have achieved their proper size and shape. Deforming forces such as wind, animals, or snow are usually detrimental. If mature organs had collenchyma for support, they would be reshaped constantly by storms or animals, which of course would not be optimal. For example, while growing and elongating, a young leaf must be supported by collenchyma if it is to continue to grow, but after it has achieved its mature size and shape, some cells of the leaf can mature into sclerenchyma and provide elastic support that maintains the leaf’s shape. Unlike collenchyma, sclerenchyma supports the plant by its strength alone; if sclerenchyma-rich stems are allowed to wilt, they remain upright and do not droop.
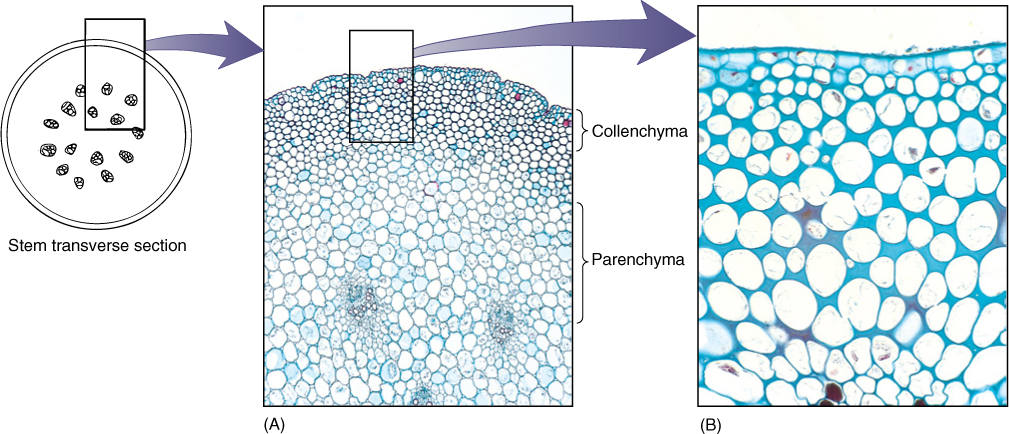
FIGURE 5-8 (A) Masses of collenchyma cells often occur in the outer parts of stems and leaf stalks. This is part of a Peperomia stem. Collenchyma forms a band about 8 to 12 cells thick. The inner part of the stem is mostly parenchyma (×50). (B) In collenchyma cells, the primary wall is thicker at the corners so the protoplast becomes rounded. No intercellular spaces are present (×150).

FIGURE 5-9 Shoot tips of long vines need the plastic support of collenchyma while their stems are elongating.
Parenchyma and collenchyma cells can absorb water so powerfully that they swell and stretch their walls, thereby growing; sclerenchyma cell walls are strong enough to prevent the protoplast from expanding. The rigidity of sclerenchyma makes it unusable for growing shoot tips because it would prevent further shoot elongation.
Sclerenchyma cells are of two types: conducting sclerenchyma and mechanical sclerenchyma. The latter type is subdivided into long fibers (Figures 5-10A to 5-10C) and short sclereids (Figures 5-10D to 5-10F and TABLE 5-3), both of which have elastic secondary walls. Because fibers are long, they are flexible and are most often found in areas where strength and flexibility are important. The wood of most flowering plants contains abundant fibers, and their strength supports the tree while their elasticity allows the trunk and branches to sway in the wind without breaking (usually) or becoming permanently bent (FIGURE 5-11). Our ribs are similarly strong, flexible, and elastic. Fiber-rich bark resists insects, fungi, and other pests.
TABLE 5-3 Types of Sclerenchyma
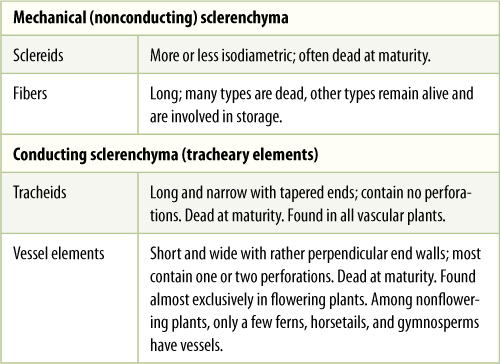
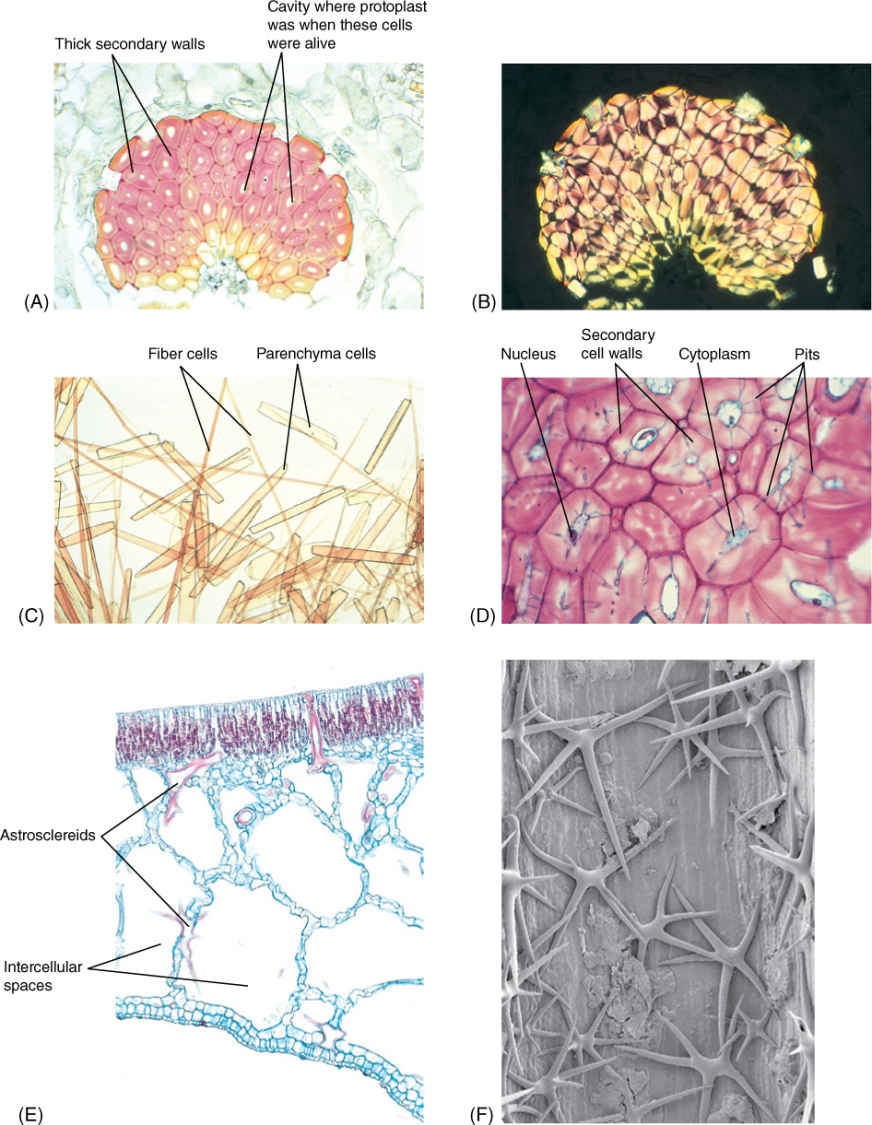
FIGURE 5-10 (A) A mass of fiber cells in the leaf of Agave. These are large, heavy, tough leaves, and fiber masses provide elastic strength. Each cell consists mostly of thick secondary cell wall; the small white space in each is an area where the protoplast had been before it died (3150). (B) The same mass of fibers as in (A) but viewed with polarized light. Secondary walls shine brightly because their cellulose molecules are packed in a tight, crystalline form, giving the wall extra strength (×150). (C) A stem of bamboo was treated with nitric and chromic acid to dissolve the middle lamellas and allow the cells to separate from each other. Fibers are long and narrow; shorter, wider cells are parenchyma (×80). (D) Sclereids are more or less cuboidal, definitely not long like fibers. These have remained alive at maturity, and nuclei and cytoplasm are visible in several. Blue-stained channels that cross the walls are pits with cytoplasm. Pits of each cell connect with those of surrounding cells so that nutrients can be transferred from cell to cell, keeping them alive (×150). (E) This portion of a water lily leaf contains large, irregularly branched cells stained red. These are astrosclereids (star-shaped sclereids). The large white spaces are giant intercellular spaces; this is an aerenchyma type of parenchyma (×40). (F) A star-shaped sclereid in a water lily leaf shown by scanning electron microscopy. These cells are located in a large intercellular space. Many have additional arms that extend back into other, smaller intercellular spaces (×200).
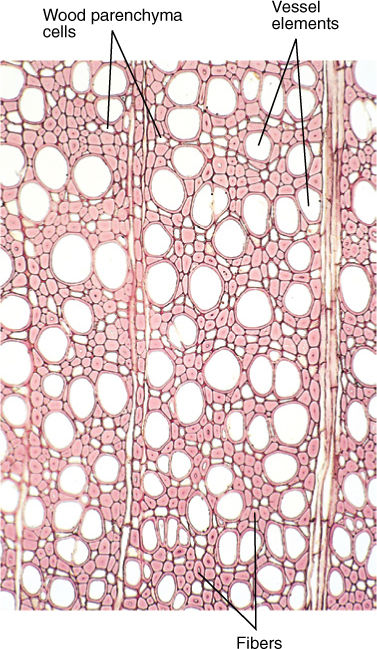
FIGURE 5-11 Wood is composed of several types of cells. The numerous small cells with thick walls and extremely narrow lumens are fibers, which give wood strength and flexibility. The large round cells that appear to be empty are vessel elements, discussed later in this chapter; the small cells with thin walls and large lumens are wood parenchyma cells (×60).
Sclereids are short and more or less isodiametric (cuboidal). Because sclereids have strong walls oriented in all three dimensions, sclerenchyma tissue composed of sclereids acts brittle and inflexible. Masses of sclereids form hard, impenetrable surfaces such as shells of walnuts and coconuts or “pits” or “stones” of cherries and peaches. Flexibility there would be disadvantageous because the soft seed inside might be crushed even though the shell remained unbroken. This is similar to our fragile brain being protected by our strong, inflexible, elastic skull.
When strength or resistance is the only selective advantage of sclerenchyma, the protoplast usually dies after the secondary wall has been deposited: A mature coconut shell is protective whether its cells are alive or dead. But in some species, certain sclerenchyma cells, especially fibers, remain alive at maturity and carry out an active metabolism (FIGURE 5-12). These living sclerenchyma cells most often are involved in storing starch or crystals of calcium oxalate. Some have rather thin secondary walls, but in others, the secondary walls are just as thick as those of fibers that die at maturity and provide only support.
Like all cells, sclereids and fibers develop from cells produced by cell division; when newly formed, they are small and have only a primary wall—they are parenchyma cells. If the cell is to differentiate into a sclereid, it may expand only slightly, but if it is to develop into a fiber, it elongates greatly. When immature sclereids and fibers reach their final size, cellulose-synthesizing rosettes in the plasma membrane begin to deposit the secondary wall. The secondary wall is located interior to the primary wall; as it becomes thicker, the protoplast must shrink, usually by removing water from its central vacuole. Secondary walls become impregnated with lignin, making them waterproof, so nutrients can enter the cell only through plasmodesmata rather than everywhere, as with parenchyma and collenchyma cells. It is important that the secondary wall not be deposited along the entire inner surface of the primary wall; small, plasmodesmata-rich areas must remain free of the secondary wall. At first, these areas are low depressions in the developing secondary wall, but as wall deposition continues, these areas become narrow pits in the secondary wall (see Figures 5-10D, 5-31, and 5-32A). The pits of adjacent sclerenchyma cells must meet; two pits make up a pit-pair. If the pits of one cell met areas of secondary wall in the neighboring cells, no water or sugars could be transferred and the protoplasts would starve.
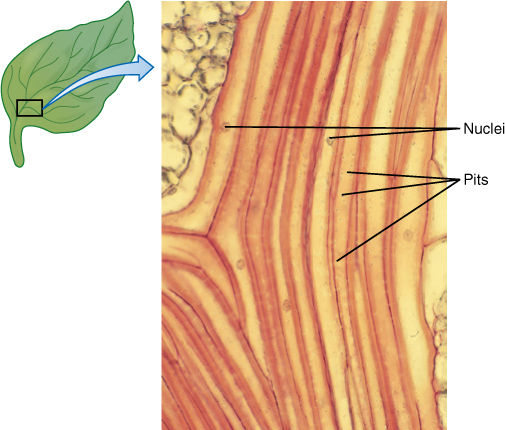
FIGURE 5-12 These fiber cells have nuclei: They are living cells. Their secondary walls are thick, but not so thick as those shown in Figure 5-11. The small dots in the walls are pits; these are much narrower than the pits shown in Figure 5-10D. Leaf of Smilax (× 150).
Conducting sclerenchyma transports water and is one of the types of vascular cells; it is discussed later in this chapter.
![]() External Organization of Stems
External Organization of Stems
The terms “stem” and “shoot” are sometimes used interchangeably, but technically, the stem is an axis, whereas the shoot is the stem plus any leaves, flowers, or buds that may be present. All flowering plants have the same basic stem organization: There are nodes where leaves are attached, and internodes, the regions between nodes (see Figure 5-1). The stem area just above the point where a leaf attaches is the leaf axil. Within it is an axillary bud, a miniature shoot with a dormant apical meristem and several young leaves (FIGURE 5-13); it is either a vegetative bud if it will grow into a branch or a floral bud if it will grow into a flower or group of flowers. The bud is covered by small, corky, waxy bud scales (modified leaves) that protect the delicate organs inside. At the extreme tip of each stem is a terminal bud. In winter, when all leaves have abscised, leaf scars occur where leaves were attached (FIGURE 5-14).
Plants and People
BOX 5-1 Parenchyma, Sclerenchyma, and Food
Most of the fruits, vegetables, and other plant parts we eat consist of almost pure parenchyma. With their thin walls, parenchyma cells are soft and easy to chew whether fresh or cooked, and they can be ground, mashed, and sliced for processed food. Whereas collenchyma and sclerenchyma are used for strength, parenchyma cells are the sites for synthesis and storage of an amazing variety of organic compounds: carbohydrates, fats, proteins, vitamins, pigments, flavors, and other nutritious materials essential to our health.
Parenchymatous foods are easy to recognize because we can bite through them easily: apples, pears, strawberries, blueberries, potatoes, lettuce, spinach, and so on. All seeds such as popcorn, beans, rice, and wheat are parenchyma as well, but in their dry, ungerminated, uncooked condition they are too hard to be edible, despite having thin cell walls. Boiling allows water to loosen hard starch grains and protein bodies and to soften the walls.
Collenchyma is not present in all plants and is never really abundant. It is almost always just a minor fraction of all the cells in leaves and stems and virtually never occurs in roots. The most familiar collenchymatous food is celery: We eat the petioles of leaves, and each ridge along the surface is a mass of collenchyma cells. Each vascular bundle inside the petiole also has a cap of collenchyma along one side. These bundles cause celery’s stringiness. Other leafy vegetables with thick petioles, like rhubarb and bok choy, also have abundant collenchyma. Most of us eat these for their flavor and nutrients, not for the pleasure of chewing endlessly on collenchyma strands.
Plants can store starch and proteins in fibers, but they do so only rarely. With the low nutrition and difficulty in chewing, we never use truly fibrous material like mature bamboo shoots and wood for food. However, many vascular bundles contain fiber cells, so even when eating parenchymatous foods we still consume some sclerenchyma. The fibers may be very noticeable, as in asparagus that is a bit too old, green beans, snow peas, artichokes, pineapples, and mangoes. In others, the fibers are a bit softer, especially after being cooked, but you may still notice them in things like squash, pumpkin, and zucchini. You have probably noticed when carving jack-o-lanterns that cleaning a pumpkin is really just a matter of removing seeds and fibers.
Sclereids also occur in some of our foods. Clusters of them cause the grittiness in pears. The seed coats of beans, peas, and most other seeds are also made up of sclereids, as is the covering on seeds of corn. The pieces that get stuck in our teeth while eating corn on the cob are composed of sclereids (and the inedible cob is mostly fibers). In small seeds that are ground into flour (wheat, rye, barley), the sclereid-rich seed coats are broken down into small pieces (the bran) that are easy to eat. Sclereids also make up the brown covering of unpopped popcorn, and popping breaks up the covering. In all cases we do not digest any part of the secondary walls of sclerenchyma cells; all the walls simply pass right through us.
The arrangement of leaves on the stem, called phyllotaxy, is important in positioning leaves so that they do not shade each other (FIGURE 5-15). If only one leaf is present at each node, the stem has alternate phyllotaxy (the leaves alternate up the stem); two leaves per node is opposite phyllotaxy, and three or more per node is whorled. The orientation of leaves at one node with respect to those at neighboring nodes is also important. In distichous phyllotaxy, leaves are arranged in only two (di-) rows (-stichies), as in corn and irises. In decussate phyllotaxy, leaves are arranged in four rows; this occurs in only some species with opposite leaves. Finally, in spiral phyllotaxy, each leaf is located slightly to the side of the ones immediately above and below it, and leaves form a spiral up the stem. This is the most common arrangement and may involve alternate, opposite, or whorled leaves (TABLE 5-4).
TABLE 5-4 Phyllotaxy


FIGURE 5-13 (A) In these buds of pistachio, leaves are modified into bud scales, just as those of prickly pear buds are modified into spines. Bud scales are waterproof and protective, but they are shed when the bud begins to grow in the spring. There are buds in the axils of the leaves and a bud at the very tip of the shoot. B) The two axillary buds of this Viburnum have already begun to grow into branches, and their first young, expanding leaves are visible. These buds were formed in the spring and started growing immediately; they did not form dormant buds with bud scales.
All flowering plant shoots are based on this simple arrangement of nodes and internodes, and diversity and specialization are variations of this arrangement. In vines, internodes are especially long, whereas in lettuce, cabbage, and onions, internodes are so short that leaves are packed together (FIGURE 5-16). Internodes can be wide (asparagus), intermediate, or narrow (alfalfa sprouts). The diversity is not random—different types provide particular adaptive advantages in certain situations. For example, plants are physically bound to the site where they happen to germinate as a seed, which by accident may be a shady area near a more optimal sunny spot. Vines, with their elongated internodes, are a means by which a plant can “explore” its immediate surroundings, and shoots that happen to grow into a sunnier site may flourish (FIGURE 5-17). In some species of climbing vines, support and attachment are provided by tendrils—modified leaves or lateral branches capable of twining around small objects.
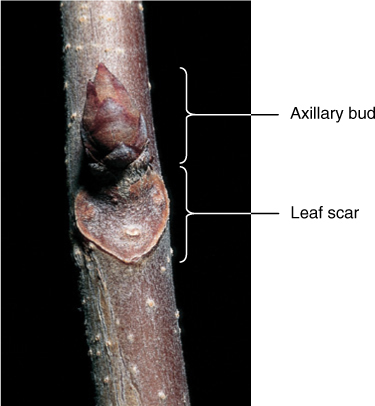
FIGURE 5-14 As a leaf falls from a stem, it breaks cells and creates a wound; a leaf scar is a layer of cork that seals the wound, keeping fungi and bacteria out and preventing water loss.

FIGURE 5-15 Phyllotaxy. (A) These young leaves of redbud (Cercis canadensis) are arranged with alternate phyllotaxy—one leaf per node. Each higher leaf is smaller and younger. (B) Leaves of Salvia show opposite phyllotaxy—two leaves per node, each pair pointing 90 degrees away from previous and subsequent pairs. (C) Irises (Iris) have distichous phyllotaxy—one leaf per node, arranged in just two rows. Iris leaves are flattened from side to side, not from top to bottom.
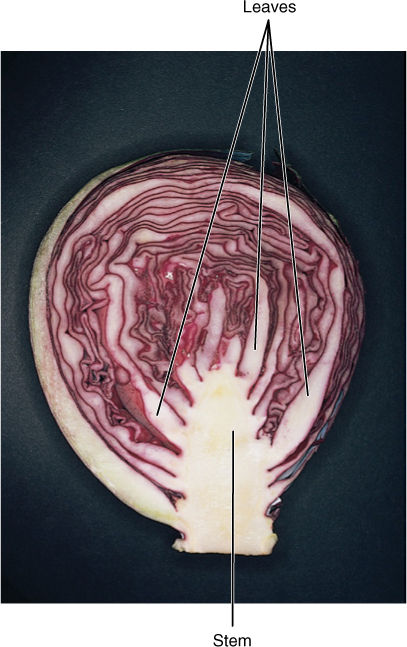
FIGURE 5-16 Cabbage nodes are packed closely together, and all leaves are tightly clustered. Such closely packed, large leaves with spiral phyllotaxy are poor at photosynthesis and did not evolve by natural selection; they were produced by plant geneticists.
The capacity to explore is even more advanced in stolons, also called runners (FIGURE 5-18). Their internodes are especially long and thin, and their leaves do not expand; thus, stolons extend greatly without using much of the plant’s nutrient reserves. After the stolon encounters a suitable microhabitat, subsequent growth is by shorter, vertical internodes and fully expanded leaves; new roots are established, and the end of the stolon resembles a new plant. If older parts of the plant die, these vertical shoots become independent plants. After they have started growing vigorously, these plants send out stolons of their own and explore further.
In some shoots, nutrient storage is particularly important for survival; these shoots are often massive and quite fleshy, thus providing many parenchyma cells in which starch accumulates. Bulbs are short shoots that have thick, fleshy leaves (onions, daffodils, garlic), whereas corms are vertical, thick stems that have thin, papery leaves (crocus and gladiolus; FIGURE 5-19). There is no obvious selective advantage of one over the other; the type seems to depend on whether mutations affecting the stem or leaf happen to occur first. Rhizomes are fleshy horizontal stems that allow a plant to spread underground (bamboo, irises, and canna lilies). Tubers are horizontal like rhizomes, but they grow for only a short period and are mainly a means of storing nutrients (potatoes). The word “tuber” is often used informally for any bulky underground plant organ.

FIGURE 5-17 (A) These grape vines (Vitis) place their leaves in the full sun at the top of the forest canopy, even though they have not invested much sugar in building strong trunks. The pines that support these vines may ultimately die because they are so heavily shaded by the grape leaves. (B) Various vines climb by different methods: This Cocculus grows in a spiral pattern and twines around slender supports. (C) Coral vine (Antigonon leptopus) climbs by means of branches that bend in a zigzag at each node, but then finally produce a tendril that wraps around objects. After the tendril attaches, the joints bend and draw the vine closer to the support. Before the tendrils attached, the jointed branch was almost straight.

FIGURE 5-18 Airplane plant (Chlorophytum) is a popular ornamental plant that spreads by runners in nature. Most people are familiar with it as a hanging basket plant, with its runners arching out and then drooping down and forming new plants at the tips. The new plant here has already produced roots.
All of these storage shoots are subterranean, which is more advantageous than an exposed surface location. Only perennial plants that go through a dormant period need storage capacity. Dormancy is the means by which perennial plants of harsh climates survive the stress of winter cold or summer heat and dryness; they often shed their leaves, reducing water loss. In order to produce new twigs, leaves, and roots in the favorable season, they must draw on stored carbohydrates. Protection of their nutrient reserves—corms, bulbs, rhizomes, and tubers—is most easily accomplished by burying them at depths that do not freeze or dry out. A subterranean location also hides them from most herbivores.
Each of these specialized tasks is accomplished by characteristic modifications in stem structure and metabolism. As with multicellularity, specialization is accompanied by division of labor: Modifications that increase a stem’s ability to survive, spread, or store nutrients decrease its efficiency at other tasks. No plant consists only of rhizomes, which are not exposed to light, because most plants must photosynthesize. At least some of the rhizome’s axillary buds must grow upward above ground and have green leaves (FIGURE 5-20). Similarly, a plant cannot be made up only of stolons: Modifications that allow quick, low-cost exploration are not appropriate for growth, photosynthesis, and sexual reproduction (flowering). Another example are the cactus-like spurges (Euphorbia) of African deserts: Each has a thick, succulent green shoot that photosynthesizes, stores water, and grows longer each year, whereas its axillary buds grow out as slender, short red branches that become fibrous then die and function as spines (Figure 5-20B). It is common for individual plants to have several types of stems and leaves, each of which contributes uniquely to the plant’s survival.
Although the axil of every leaf contains a bud, only a few buds ever develop into a branch; the others remain dormant or produce flowers. Axillary buds do not become active at random; each species has a particular pattern. In shady environments, it may be advantageous for axillary buds to remain dormant so that all resources are concentrated in the growth of the vertical main shoot, the trunk, allowing the plant to reach brighter light in the top of the forest canopy (FIGURE 5-21). In an open environment, such dominant upward growth is not particularly advantageous; it may be better for all buds to grow and thereby maximize the rate at which new leaves are formed. On almost all plants, at least a few axillary buds remain dormant and serve as reserve growth centers. As long as the apical meristem is healthy and growing well, some axillary buds are unneeded; if the apical meristem is killed by frost, insects, or pruning, axillary buds become active and replace it, allowing the growth of the shoot to continue.
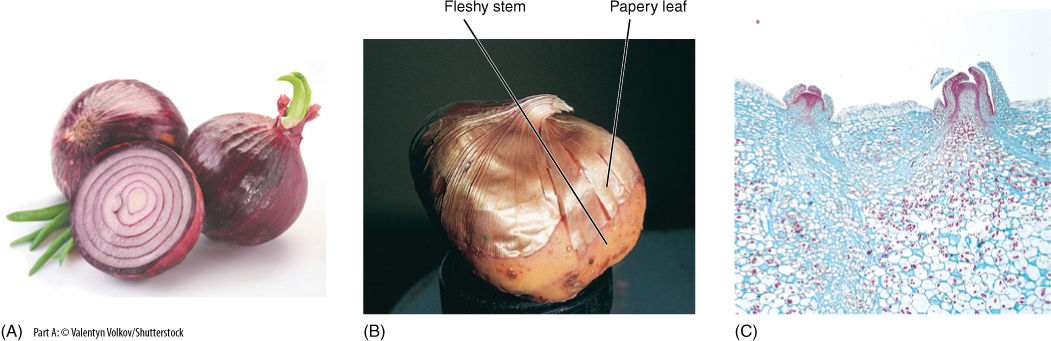
FIGURE 5-19 (A) Onions are bulbs: They have short vertical stems and fleshy leaves. (B) Gladioluses have corms with fleshy stems and papery leaves. (C) A tuber (potato: Solanum tuberosum) is a short horizontal stem that grows only for a limited period of time. Its leaves are microscopic, and its axillary buds are the potato’s “eyes,” shown here by light microscopy (× 15).

FIGURE 5-20 (A) Division of labor. False Solomon’s seal (Smilacena racemosa) consists of subterranean rhizomes (which survive harsh winters and spread through the soil) and aerial shoots (which carry out photosynthesis and flowering). (B) This succulent spurge (Euphorbia) has two types of shoots: the long wide green shoot that lives for many years, and many narrow, red shoots that die quickly then function as spines.
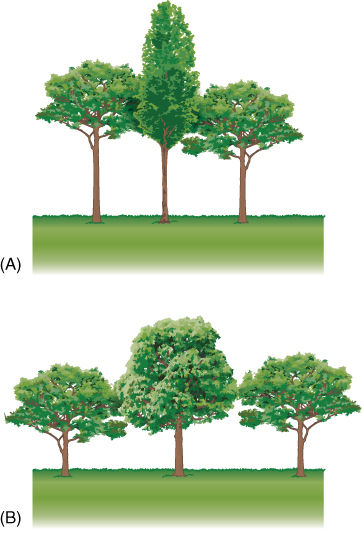
FIGURE 5-21 (A) In a heavily shaded forest, many axillary buds remain permanently dormant, allowing all resources to be concentrated on placing one shoot up into the light. (B) With abundant light, one main trunk is not so advantageous; instead, many axillary buds grow as branches, allowing rapid production of leaves.
![]() Internal Organization of Stems: Arrangement of Primary Tissues
Internal Organization of Stems: Arrangement of Primary Tissues
In order to function properly, a tissue must contain the right cells in the proper arrangement. The same principle is true on a larger scale: In order to function properly, the tissues of an organ must be arranged correctly.
Epidermis
The outermost surface of an herbaceous stem is the epidermis, a single layer of living parenchyma cells (FIGURE 5-22). This differs greatly from our human epidermis, which is much thicker, consisting of many layers of dead cells.
Also, whereas we humans have lungs and a digestive tract that absorbs certain things and expels others, all interchange of material between a plant and its environment occurs by means of its epidermis.
Because air is almost always drier than living cells, even when relative humidity is high, preventing loss of water to air is a critical function for the epidermis of land plants. Also, the epidermis is a barrier against invasion by bacteria and fungi as well as small insects. It shields delicate internal cells from abrasion by dust particles, passing animals, or leaves and stems that might rub together, and its reflectivity protects the plant from overheating in bright sunlight. Although it might seem that thick-walled sclerenchyma cells would be more protective, epidermis cells are almost always thin-walled parenchyma cells.
Plants Do Things Differently
BOX 5-2 Organs: Replace Them or Reuse Them?
Plants have three types of organs: roots, stems, and leaves. Animals have many types: heart, brain, liver, eyes, kidneys, urinary bladder, and so on. Notice that we have only one or two of each: one heart, one brain, two eyes. Plants, on the other hand, typically have many of each: dozens of stems, hundreds of roots, and thousands of leaves. Also, notice how long each lasts relative to the life of the organism. We have only one heart during our entire lifetime. If it fails, we die. Considering all the mechanical work our heart must do, it is remarkable that we do not periodically grow a new one. We have two kidneys, so if one stops functioning, we have a backup organ; however, our bodies will not grow a replacement, even if we lose both kidneys. If we are lucky enough to die of old age, the organs we have at that time are the same ones we were born with.
Plants are very different. Their biology is based on using an organ just once then throwing it away and replacing it with a new version. A tree might have thousands of leaves, but it will discard those in autumn and make an entire new set the following spring. When the tree blooms, it might have thousands of flowers, each with many petals and other parts. The petals function for just a day or two then wither and fall off. If the flower was pollinated, it will form a fruit, and then that too will fall off. After the flowering period is finished, the plant will have no sex organs whatsoever until the following year, when it makes a whole new set. Think about how different our biology is from that. Our first thought is probably relief that our sex organs do not fall off after use, even if we could grow another set next year.
What are the consequences of the plant-type alternative of depending on “one-use” organs versus the animaltype of “reusable” organs? Leaves must be exposed to sunlight if photosynthesis is to occur, so they suffer extensive exposure to ultraviolet light. Ultraviolet light causes mutations in both plants and animals, leading to skin cancer in us and to damaged, nonfunctional leaves in plants. We avoid ultraviolet light by wearing clothing (other land animals have hair, feathers, or scales) or by performing our activities in shade or indoors. But leaves must be in light. In addition, leaves become covered with dust, spores, fungi, bacteria, insect poop, and other debris. They become just as dirty as any animal, but leaves cannot groom themselves. By abscising their leaves and growing a new set, they get rid of dirty, diseased, broken-down organs and start off a new year with fresh, clean ones. If plants had to rely on the same leaves for their entire lifetime, no tree would ever make it to 100 years old.
The same principle applies to other one-use plant organs. Some trees live to be not just hundreds of years old but thousands. Even at such an amazing age, whenever the tree undergoes sexual reproduction, it makes brand new flowers (or cones), and those are as fresh and vigorous as when the tree was only a few years old. We animals are not so lucky. When we get on in years, we may still be willing, but our reproductive organs are old. In men, sperm cells are produced by division of the same set of cells that became active when we went through puberty, and there is no reason to think that those cells will remain in better shape than those of our skin (wrinkled), eyes (failing), hair follicles (bald long ago), teeth (false). In women, cells that will mature into eggs all begin meiosis before we are even born and have been in cell cycle arrest ever since, with just one egg maturing each month. By the time we are 40 or 50 years old, so are all our potential egg cells—but not in plants, not even in old plants. All of their eggs and sperm cells are brand new made in flowers that have just been produced only a week or two before.
Even our circulatory systems follow this difference. We have just one set of blood vessels all our lives. At this point, your aorta is about 20 years old, as are the arteries that take blood up to your brain. All the blood that goes to and from your lungs is passing through veins about one fifth of a century old, and they are getting older as you read this. Not in plants, however. All woody plants make an entirely new vascular system every year. Every springtime, plants make a new layer of wood that conducts water and a new layer of bark that conducts sugars and minerals. The wood and bark from last year may still be functional, or it may be damaged by fungus; however, that does not matter. There is a set of new tissue, as healthy and functional as our human tissues were when we were babies.
Plants and animals are not totally different, actually. We threw away our baby teeth and replaced them with a set of permanent teeth when we were about 6 years old—a very plant-like thing to do. And sharks do not stop with just two sets—they constantly produce replacement teeth as long as they live. Birds replace their feathers every year, and starfish grow a new arm if one is cut off. Some lizards grow a new tail, but such capacity does not occur in us: Other than baby teeth, our organs must last us a lifetime. But any time you look at a plant, you are seeing only a tiny fraction of the plant. You are not seeing all the parts that have already existed and fallen off nor all the parts that the plant will have in the future. If all those parts could be gathered up and preserved, the total plant would be vastly greater than it ever is at any particular time.
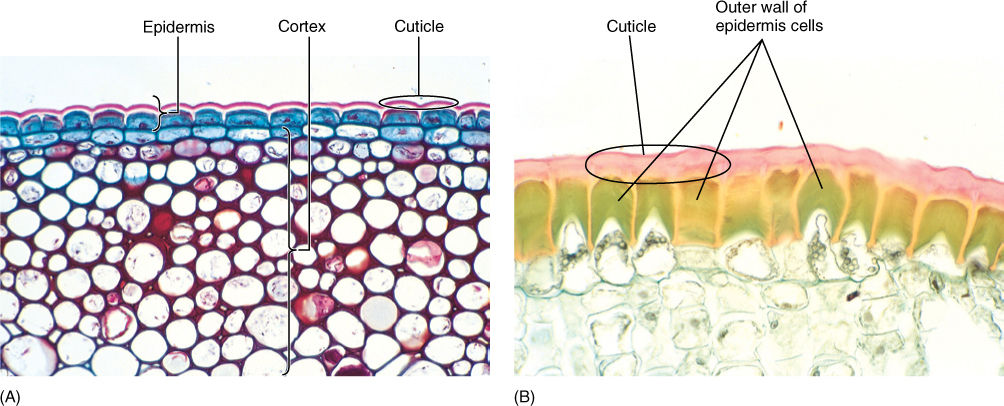
FIGURE 5-22 (A) The cuticle on this ivy (Hedera helix) has been stained dark pink; the lighter layer is wall material impregnated with cutin. Epidermal protoplasts have stained dark blue. Interior cells are photosynthetic collenchyma (× 120). (B) This epidermis was taken from a desert plant, Agave. Its cuticle is much thicker than that of ivy in (A), and the outer walls of the epidermal cells are extremely thick. Both the wall and cuticle strengthen the leaf surface against insects and prevent water loss (× 200).
The outer tangential walls are encrusted with cutin, a fatty substance that makes the wall impermeable to water (Figure 5-22). In species that occur in mild, moist habitats, such encrustation may be sufficient for water retention, but in most plants, cutin builds up as a more or less pure layer called the cuticle. Under more severe conditions, even a cuticle is not sufficient, and a layer of wax may be present outside the cuticle (FIGURE 5-23 and TABLE 5-5). Cutin and wax resist digestive enzymes and provide defense against pathogens like fungi and bacteria. In some species, the cuticle is so smooth that fungal spores cannot even stick to it and are washed off by rain or shaken off by wind. Waxes are indigestible and nonnutritious, therefore, a thick layer of wax makes it difficult and unrewarding for an insect to chew into a stem.

FIGURE 5-23 On this epidermis, wax is present as flat plates, but in other species it can occur as threads, beads, flakes, or liquid (× 550).
Unfortunately, cutin and wax also inhibit the entry of carbon dioxide needed for photosynthesis—a totally impermeable epidermis would lead to a plant’s starvation. However, the epidermis contains pairs of cells (guard cells) with a hole (stomatal pore) between them (FIGURE 5-24). Guard cells and a stomatal pore together constitute a stoma (plural, stomata). Stomatal pores can be opened during the daytime, permitting carbon dioxide to enter the plant.
The opening of the stomatal pore is made possible by the structure of the guard cells. Guard cell walls have a special radial arrangement of cellulose microfibrils, which causes some parts to be weaker and more extensible than others. Guard cells swell by absorbing water; the walls next to the stomatal pore are rigid and do not extend, but the back walls are weak and stretch (FIGURE 5-25), causing the cells to form an arc and push apart, opening the pore and allowing entry of carbon dioxide and exit of oxygen. This unavoidably allows water vapor to escape and also increases the risk of microbes entering the plant. After sunset, when photosynthesis is impossible, it is not advantageous to keep stomata open; guard cells in most species shrink and straighten, closing the pore and preventing water loss.
TABLE 5-5 Special Chemicals in Walls
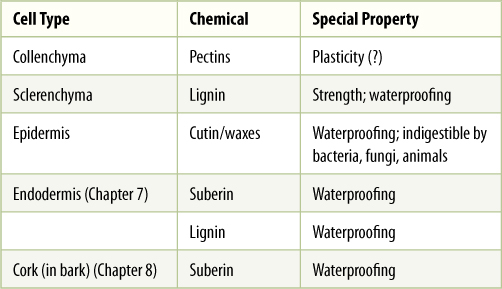
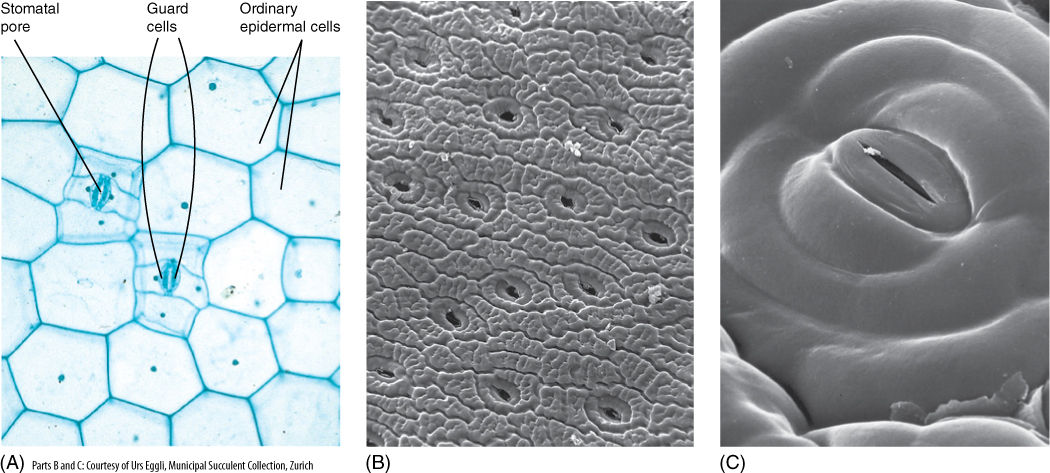
FIGURE 5-24 (A) Light microscope view of the epidermis of Zebrina; the stomatal pore, guard cells, and adjacent cells are visible, as are ordinary epidermal cells (×150). (B) Scanning electron micrograph of the epidermis of a cactus showing numerous stomata. Notice how close together stomata are; no chlorenchyma cell inside the cactus (on the other side of the epidermis) is far from a point where carbon dioxide can enter (×50). (C) The stomatal pore is surrounded by guard cells and then other epidermal cells. Notice the small particle of debris at one end of the pore; clogging by debris is a problem in dusty areas (×800).
In most plants, some epidermal cells elongate outward and become trichomes, also called hairs (FIGURE 5-26). Trichomes make it difficult for an animal to land on, walk on, or chew into a leaf. They shade underlying tissues by blocking some incoming sunlight, which may be too intense in summer or in high alpine environments. Trichomes can also create a layer of immobile air next to a leaf surface, which allows water molecules that diffuse out of a stoma to bounce back in rather than be swept away by air currents.
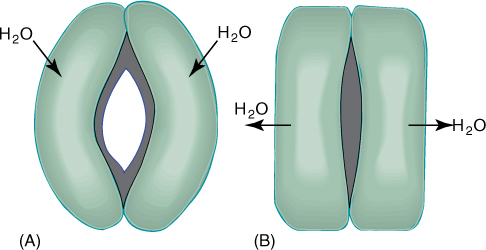
FIGURE 5-25 (A) When guard cells absorb water and swell, they become curved and the inner walls are pulled apart by the expansion of the back walls. (B) If guard cells lose water and shrink, they straighten, closing the stomatal pore.
Trichomes exist in hundreds of sizes and shapes; many are unicellular, being just long, narrow epidermal cells. Multicellular ones may be a single row of cells or several cells wide, and many branch in elaborate patterns. Most trichomes die shortly after maturity, and their cell walls provide protection; but others remain alive and act as small secretory glands. Some secrete excess salt; others produce antiherbivore compounds, and those in carnivorous plants secrete digestive enzymes onto trapped insects. The poisonous, irritating compounds of stinging nettle are held in trichomes.
Cortex
Interior to the epidermis is the cortex (FIGURE 5-27). In many plants, it is quite simple and homogeneous, composed of photosynthetic parenchyma and sometimes collenchyma. In other species, it can be very complex, containing many specialized cells that secrete latex, mucilage, or pitch (resin). Some cortex cells contain large crystals of calcium oxalate or deposits of silica.
Cortex cells of most plants fit together compactly, but in fleshy stems such as tubers, corms, and succulents, cortex parenchyma is aerenchyma, an open tissue with large intercellular air spaces (see Figures 5-6 and 5-10). A few angiosperms, such as water lilies and the plants grown in aquaria, have become aquatic, living submerged in lakes or oceans. These plants have large cortical air chambers that provide buoyancy. The stems have such a strong tendency to float that no sclerenchyma is necessary for support.
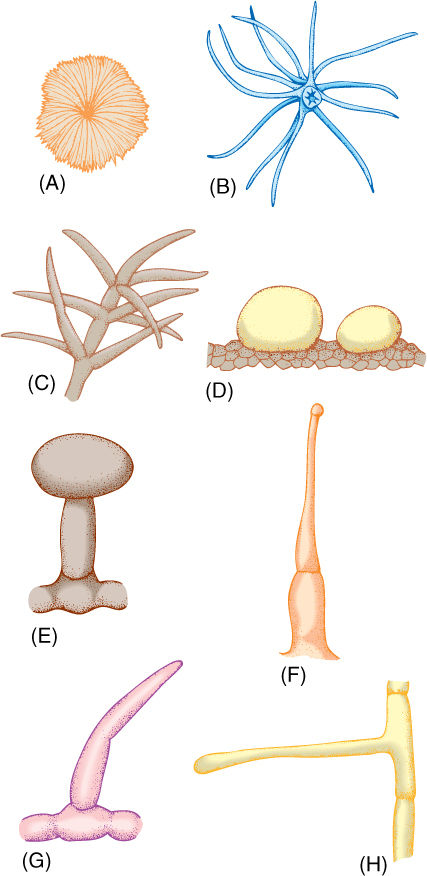
FIGURE 5-26 Trichomes. (A) Flat, scale-shaped trichome. (B and C) Branched trichomes. (D) Glandular trichomes—these are two large single cells. (E) Glandular trichome with a stalk cell elevating the secretory head cell. (F) Glandular trichome, capable of injecting poison. (G and H) Simple, unbranched, nonglandular trichomes.
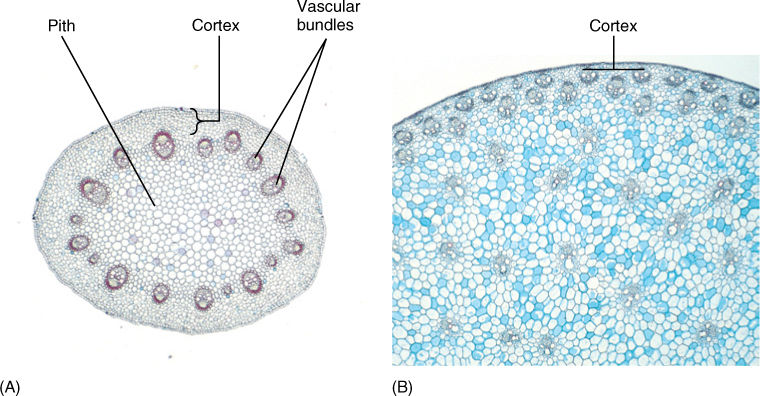
FIGURE 5-27 (A) The cortex of this stem of buttercup (Ranunculus) is the narrow band of cells between the epidermis and the vascular bundles (×20). (B) The cortex of this corn stem is even narrower, in some places only two or three cells wide (×20).
Vascular Tissues
For very small organisms whose bodies are either unicellular or just thin sheets of cells, diffusion is adequate for the distribution of sugars, minerals, oxygen, and carbon dioxide throughout the body (Box 5-3). But if any cells of the organism are separated from environmental nutrients and oxygen by just four or five cell layers, diffusion is too slow, and a vascular system is necessary. Two types of vascular tissues occur in plants: xylem (pronounced ZY lem), which conducts water and minerals, and phloem (pronounced either FLOW em or flome), which distributes sugars and minerals.
Vascular systems of plants are quite different from those of animals. The plant vascular system is not a circulatory system: Water and minerals enter xylem in the roots and are conducted upward to leaves and stems. During its passage, xylem sap travels through dead, hollow cells, not through tubes composed of living cells like our blood vessels. Once in the shoots, most water evaporates from the surfaces of stems, leaves, and flowers and is lost; the minerals and a bit of water are used by surrounding cells. Phloem cells are living; they pick up sugar from areas where it is abundant, usually leaves during summer and tubers or rhizomes in spring, and transport it to areas where sugar is needed, especially growing tips of shoots, roots, young leaves, and flowers. In the later months of summer, phloem carries sugar into developing fruits and into the storage organs of perennial plants. Because sugar must be dissolved to be conducted, water is transported simultaneously in phloem.
Xylem
Within xylem are two types of conducting cells: tracheids and vessel elements; both are types of sclerenchyma (FIGURE 5-28 and TABLE 5-6). The term “tracheary element” refers to either type of cell. As a young cell matures into a tracheary element, it first must enter cell cycle arrest and stop dividing. It is initially a small parenchyma cell with only a thin primary wall, but the cell becomes long and narrow and then deposits a secondary wall that reinforces the primary wall. The cell then dies, and its protoplasm degenerates, leaving a hollow tubular wall (FIGURE 5-29).
Alternatives
BOX 5-3 Simple Plants
Three groups of plants have much simpler bodies than those described in this chapter. Hornworts (Anthocerotophyta) are a rather rare group of tiny plants whose bodies look like bits of green cellophane, often no larger than 2 to 5 mm in diameter and only a few cells thick. They have no epidermis, no stomata, no collenchyma, no sclerenchyma, no cortex, no pith, no xylem, and no phloem. Instead, they are small ribbon- or disk-shaped sheets of parenchyma cells, most of which have a single chloroplast. Each cell absorbs water and minerals, carries out its own photosynthesis, and is relatively self-reliant. When the plant reproduces, horn-like structures grow upward, break open, and release spores (there are no flowers). During this stage, nutrients are transported from chlorophyllous cells to spore-producing cells, but only by means of plasmodesmata, not phloem.
Many liverworts (Hepatophyta) are about as simple as hornworts. Plants of Pallavicinia, Sphaerocarpos, and Riccia also consist of just small green ribbons or disks of chlorophyllous cells. Bodies of Fossombronia and Petallophyllum appear leafy and more complex, but in fact, they are just ribbons whose edges are puckered. The bit of complexity present in these bodies is merely that the central portion of the ribbon is several cells thick, whereas the margins are unistratose. As in hornworts, there is no epidermis or vascular tissue.
Other liverworts such as Asterella and Marchantia have thicker, more complex bodies. The ventral side (the “belly” side facing the soil) is several layers of compact parenchyma with few chloroplasts, and the dorsal side (the “backbone” side facing the air) consists of intersecting sheets of cells that form air chambers (like the intersecting sheets of cardboard that form a six-pack carton for beer). The term “epidermis” is used for surface cells; dorsal epidermis has stoma-like air pores. Ventral epidermis has trichomes (called rhizoids) that attach the body to soil or tree bark. The most complex liverworts have “leaves” and are called leafy liverworts, but their bodies are composed of just parenchyma. As in hornworts, liverworts transfer nutrients through plasmodesmata from chlorophyllous cells to spore-producing tissues when they reproduce: Liverworts have no vascular tissues. Despite their simplicity, all liverworts grow by means of organized apical meristems.
Mosses (Bryophyta) are a bit more complex than either hornworts or liverworts. Mosses always have a stem with leaves, and in those with taller stems or larger leaves, some cells deposit a thicker wall and resemble sclerenchyma fibers. However, these walls do not contain lignin and probably have a separate evolutionary origin, so they are called stereides instead of fibers to avoid confusion. Many mosses conduct water, but along the outside of their bodies, not inside and not with xylem: Their numerous leaves are so tiny and close to the stems they form capillary spaces and act like a wick. A number of mosses do have an internal system of conducting cells in their green, leafy stems; cells called hydroids conduct water, and others called leptoids conduct carbohydrates and other nutrients. These should never be called “xylem” or “phloem” because they differ in significant ways from those two tissues.
What are some consequences of having such simple bodies? The simplicity of hornworts, liverworts, and some mosses limits their ability to survive in some habitats. Lacking conducting tissues, they must remain short, thin, or prostrate, and many quickly die in dry air. On the other hand, this same simplicity enables them to survive in many habitats because just a few weeks of cool moist weather is enough to allow their spores to germinate and grow into tiny, mature plants. They do not need to photosynthesize long enough to produce all the carbohydrates that a vascular plant needs for its larger, more complex body. Even deserts have rainy periods and shaded spots that stay moist for weeks; such spots will almost always have some nonvascular plants thriving there.

FIGURE B5-3A Plants of hornworts (Phaeoceros); the bodies are just thin, green sheets of parenchyma. The columns (“horns”) are reproductive structures.

FIGURE B5-3B Plants of this liverwort (Riccia fluitans) grow in quiet streams; each shoot is less than 1 mm thick and consists of parenchyma cells. Each grows from an apical meristem that occasionally divides in two.
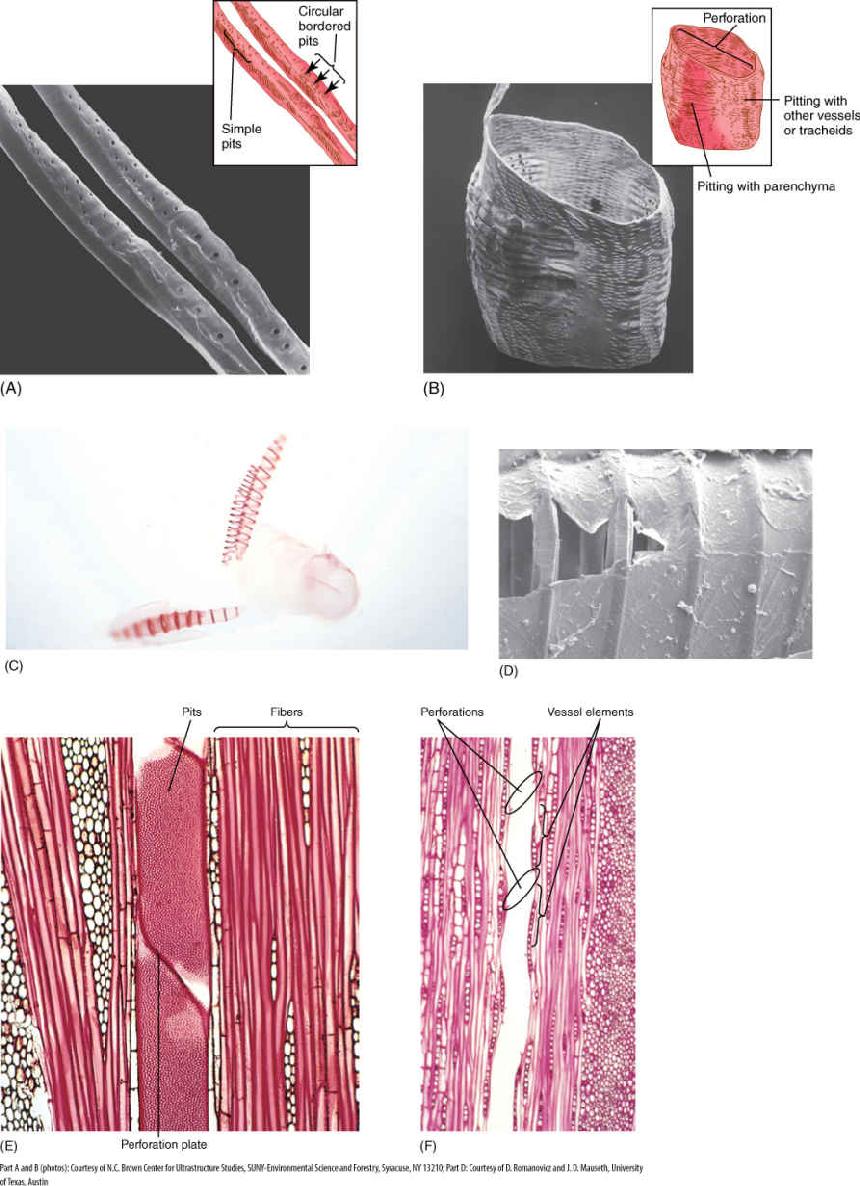
FIGURE 5-28 Tracheary elements. (A, C, and D) Tracheids are long cells with tapered ends. The primary wall is complete over the whole surface, and in (C), it is so thin that the inner, spiral secondary wall is visible. In (D), the primary wall tore during specimen preparation, revealing just how thin it is and permitting views of the circular bands of secondary wall which are located just interior to the primary wall and which prevent it from collapsing during water conduction. (B and E) Vessel elements tend to be wider and shorter than tracheids, but the most important feature is the perforation, the large hole at each end. (F) The perforation of one vessel element must be aligned with that of the next if water is to pass through with little friction; the stack of vessel elements is a vessel. The vessel here has been cut down the center, so the front wall and back wall are missing and the vessel appears to be just an open space. (A) ×600 (B) ×270 (C) ×100 (D) ×1000 (E) ×200 (F) ×50.
TABLE 5-6 Tracheary Elements (Conducting Sclerenchyma)
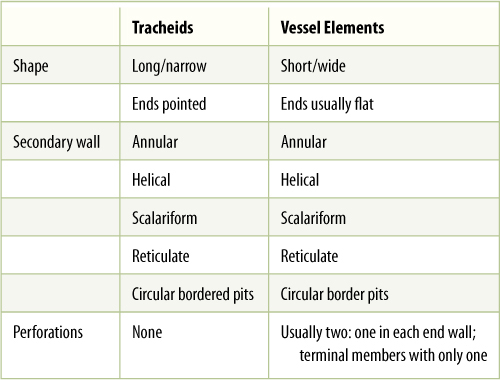
The secondary wall is impermeable to water; thus, areas of the permeable primary wall must remain uncovered if water is to enter and leave the cell. In the simplest type of tracheary element, there is only a small amount of secondary wall, organized as a set of rings, called annular thickenings, on the interior face of the primary wall (Figure 5-29E). This arrangement provides a large surface area for water movement into and out of the cell, but it does not provide much strength. The primary wall must be supported because movement of the water tends to cause it to collapse inward (FIGURE 5-30). With helical thickening, the secondary wall exists as one to three helices interior to the primary wall (Figure 5-29F). Scalariform thickening provides much more strength because the secondary wall underlies most of the inner surface of the primary wall and is fairly extensive. Just as in fibers and sclereids, the area where the secondary wall is absent is called a pit (FIGURE 5-31A). In tracheary elements with reticulate thickening, the secondary wall is deposited in the shape of a net, as the name suggests. The most derived and strongest tracheary elements are those with circular bordered pits (FIGURE 5-31B and FIGURE 5-32). In such tracheary elements, virtually all of the primary wall is underlain by secondary wall. The pits that allow water movement are weak points in the wall, but the weakness is reduced by a border of extra wall material around the pit, hence the name.
Tracheary elements with annular thickenings are weak, but a large percentage of their primary wall is free of secondary wall and available for water movement (FIGURE 5-33). Pitted tracheary elements are just the opposite: They are extremely strong, but so much of the surface is underlain by lignified secondary wall that water enters or leaves the cells slowly. Each is selectively advantageous under certain conditions and disadvantageous under others. If a species grows in perpetually wet soil and water moves easily, its cells are not in danger of collapsing, and thus, tracheary elements with annular or helical secondary walls are adaptive, not only because the large open regions of primary wall allow water to pass through quickly but also because the plant uses little sugar building secondary walls. If soil dries even slightly, however, water is pulled with more force and exerts inward traction on tracheary element walls. The stronger scalariform, reticulate, or circular bordered pitted walls are necessary to keep the tracheary elements open. Although they cannot conduct water as quickly, water movement is automatically slower because the soil is dry. Under dry conditions, using extra glucose to build stronger secondary walls can be a valuable investment.
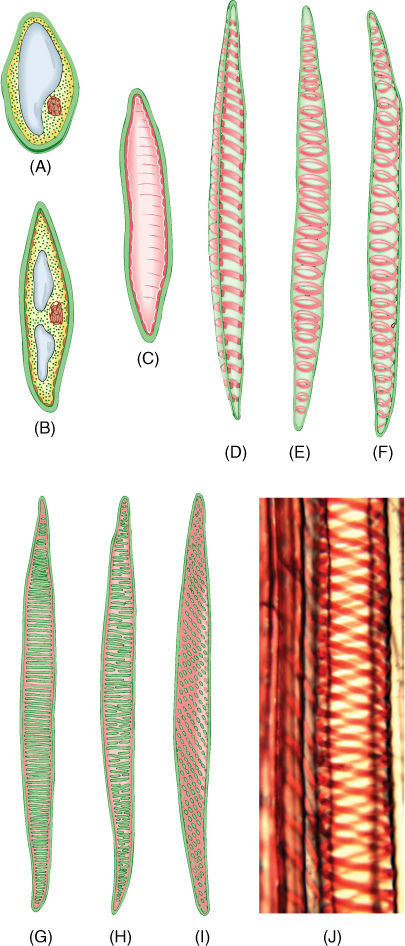
FIGURE 5-29 Tracheids. (A—D) The growth of a young cytoplasmic cell into a tracheid. After the cell has almost reached mature size, the secondary wall is deposited interior to the primary wall (C). After the secondary wall is finished, the protoplasm dies and degenerates, leaving only the primary and secondary walls. (D) A tracheid cut open, showing a secondary wall in the form of helical thickenings. (E) A whole cell in which the primary wall is so thin that the annular secondary walls inside are visible. (F) Whole cell with helical secondary wall. (G) Scalariform secondary wall. (H) Reticulate secondary wall. (I) Pitted secondary wall. (J) Light micrograph of a tracheid with a helical secondary wall (×500).
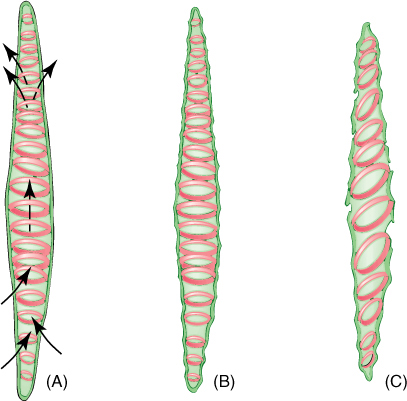
FIGURE 5-30 (A) As water enters a tracheary element, it passes through primary wall between regions of secondary wall. (B) Water adheres to cellulose molecules, pulling walls inward and upward as it rises in xylem; primary walls tend to collapse but secondary walls provide strength to hold the primary wall in place. (C) Under severe water stress (dry air and dry soil), annular and helical secondary walls may not be strong enough to prevent collapse of the primary wall.
Tracheids obtain water from other tracheids below them and pass it on to those above. They must occur in groups, lying side by side with some and having their ends overlapping the ends of others (FIGURE 5-34). Also, the pits of adjacent tracheids are aligned so that water can pass through. The aligned set of pits is a pit-pair, and the set of primary walls and middle lamella between them constitutes a pit membrane, just as in fibers and sclereids (Figure 5-31). (“Pit membrane” is an old term and should not be confused with lipid/protein membranes such as the plasma membrane.) Although very permeable to water, a pit membrane—being a set of primary walls—does offer slight resistance. If each tracheid is 1 mm long, each water molecule passing from root to leaf in even a short plant only 1 meter tall (1,000 mm) must pass through 1,000 pit membranes. The friction adds up.
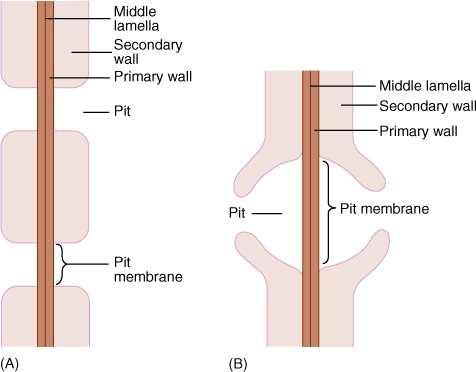
FIGURE 5-31 (A) Side view of a pit that has been bisected. In a sclereid or fiber, the secondary wall would be thicker and the pit narrower and deeper. The diameter of the pit can vary, being broad as in Figure 5-10D or narrow as in Figure 5-12. (B) A bordered pit has a rim of extra wall material that prevents the pit from being a weak spot in the wall. With borders, pits can be broad enough to allow rapid flow of water from tracheary element to tracheary element, but not so weak as to be detrimental. Only tracheary elements have bordered pits; fibers and sclereids do not.

FIGURE 5-32 (A) Bordered pits in face view on front wall of a vessel element. The white slits are the actual openings of the pits, the pinkish area around each white region is the border, and the dark red hexagons are regions where the secondary wall lies against the primary wall (×400). (B) Bordered pits in face view on the front walls of tracheids of pine. This is the same magnification as (A); notice that tracheids are much narrower than vessel elements, and pits in pine are extremely large. (C) Scalariform bordered pits (×700).
Vessel elements provide a way to move water with less friction than tracheids. Vessel elements, like tracheids, are individual cells that produce both primary and secondary walls before they die at maturity. In vessel elements, however, an entire region of both primary and secondary wall is missing. During the final stages of differentiation, a large hole called a perforation is digested through a particular site of the primary wall, often removing the entire end wall (Figures 5-28, 5-33, and 5-35). The perforations of adjacent vessel elements must be aligned, and each element must have at least two perforations, one on each end. Whereas water must pass through pit membranes each time it enters or leaves a tracheid, it passes through perforations as it goes from one vessel element to the next. Because perforations are wide and completely lack primary walls, they cause little friction.
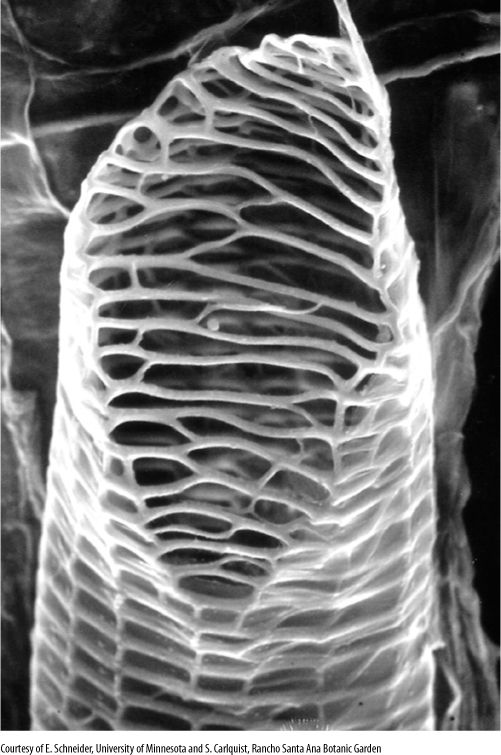
FIGURE 5-33 This vessel element was isolated from tissue by dissolving the middle lamella. The secondary wall (the network along the cell’s end and sides) is clearly visible on the end because the primary wall was digested away before the cell died; these open holes constitute a perforation. The thin, almost transparent covering on the cell’s sides is the primary wall, which was not digested. In the plant, another vessel element would have been stacked above this one, with their perforations aligned and permitting water flow (× 1,000).
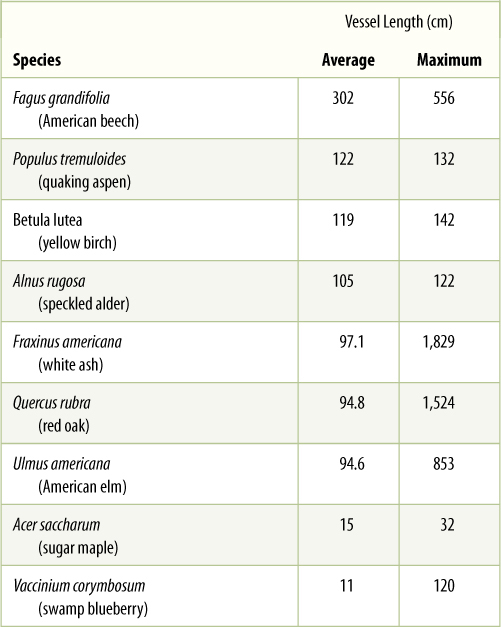
An entire stack of vessel elements is a vessel. The vessel elements on each end of a vessel have only one perforation. Whereas vessel elements, being individual cells, are only about 50 to 100 mm long, vessels can be many meters long, running all the way from a root tip to a shoot tip, although some are only a few centimeters long (TABLE 5-7).
Vessels must absorb water from parenchyma cells, tracheids, or other vessels, and they must pass it on. Their side walls have pits for this lateral transfer (Figure 5-34), and all of the types of wall thickening mentioned above for tracheids also occur in vessel elements. The only constant difference is that vessel elements have perforations—complete holes—in their walls, whereas tracheids only have pits (Table 5-6). Perforations greatly reduce friction, and water moves much more easily through a set of vessels than through a set of tracheids. Tracheids and vessel elements have plasmodesmata in their primary walls, but these are too narrow to be important in long-distance water conduction; typical diameters—plasmodesmata, 0.04 μm; pits, 1 to 2 μm; and perforations, 20 to 50 μm.
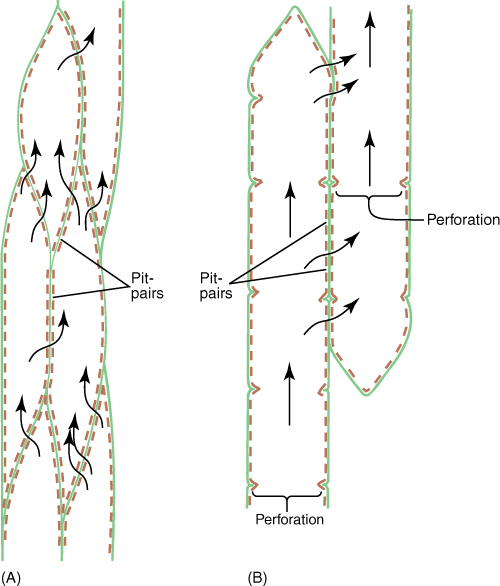
FIGURE 5-34 (A) Water passes between tracheids only through pit-pairs. Other than plasmodesmata (which are too tiny to be important in water conduction), no holes are present in primary walls of tracheids. (B) Water can pass between vessel elements through perforations, but it can pass from one vessel to another only through pit-pairs. Vessels are long, but not as long as a whole plant, and water must, therefore, pass from one vessel to another as it moves upward from roots to stems to leaves. Green = primary wall; brown = secondary wall.
Within our human circulatory system, all arteries, capillaries, and veins share the same lumen: A water molecule or a red blood cell could circulate through every part of our blood vessel system and never encounter a barrier. In contrast, water molecules must move across pit membranes every time they enter or leave a tracheid or vessel. Some consequences are that the plant system causes more friction than does our vascular system, and solid particles cannot be transported in plants. However, a hole or leak in our system could potentially let all our blood leak out (and is often fatal), whereas a hole in a plant system affects only one tracheid or vessel, not the entire system (and is never fatal).
Tracheids evolved at least 420 million years ago, and virtually all plants with vascular tissue have tracheids. Vessels evolved more recently and occur almost exclusively in flowering plants, where they perform long-distance water conduction in roots and stems. In flowering plants, tracheids occur mostly in the fine veins of leaves but can also be found in the wood of some species. Conifers such as pines and redwoods have xylem composed exclusively of tracheids. Until recently, we thought that ferns too contained only tracheids, but they can at least partially digest their pit membranes and convert pit-pairs into sets of perforations (FIGURE 5-35).
TABLE 5-7 Length of Vessels in Trunks of Trees
Phloem
Like xylem, phloem has two types of conducting cells, sieve cells and sieve tube members; the term “sieve element” refers to either (FIGURE 5-36). These are very different from tracheary elements: They are parenchyma cells, have only primary walls, and must remain alive in order to conduct. As immature sieve elements begin to differentiate, their plasmodesmata enlarge to a diameter of more than 1 mm and are called sieve pores. Plasmodesmata occur in groups (primary pit fields); therefore, sieve pores also occur clustered together in groups called sieve areas. Sieve elements remain alive during differentiation, and the plasma membrane that lined the plasmodesma continues to line the sieve pore (FIGURE 5-37). The amount of cytoplasm within the pore also increases, and rapid, bulk movement from cell to cell becomes possible. Sieve pores of adjacent cells must be aligned if phloem sap is to pass through.
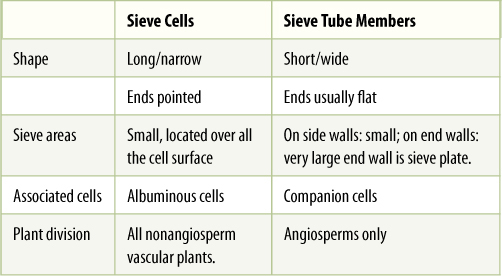
The two types of sieve elements differ in shape and placement of sieve areas (Figure 5-36). If the cell is elongate and spindle shaped (like a tracheid) and has sieve areas distributed over all its surface, it is a sieve cell (TABLE 5-8). This type evolved first and is found in older fossils and in nonangiosperm vascular plants. But because phloem conducts mostly longitudinally, it is most efficient for sieve areas on the two ends of the cell to be especially large and to have very wide sieve pores, whereas sieve areas on the sides of the cell can be rather small. Cells like this are sieve tube members; they are stacked end to end with their large sieve areas aligned, forming a sieve tube. These end-wall sieve areas with large sieve pores are sieve plates. Like vessels, sieve tubes are shorter than the plant, and the small, lateral sieve areas are important for transport of phloem sap from one sieve tube to another. Sieve tube members must have evolved at about the same time as the flower; all angiosperms have sieve tube members, but none of the nonangiosperms has them.
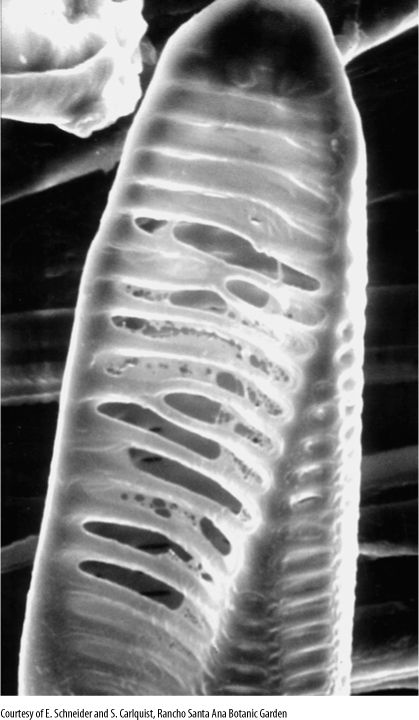
FIGURE 5-35 This fern (Astrolepis) tracheary element had digested portions of primary wall out of several pits, converting them to crude perforations. In angiosperms, the wall-digesting mechanism is more refined and removes the primary wall more completely.
An unusual feature of sieve elements is that their nuclei degenerate but the cells remain alive. Cytoplasm cannot carry on complex metabolism without a nucleus, and in phloem, the necessary nuclear control is provided by cells intimately associated with conducting cells. Sieve cells are associated with albuminous cells, and sieve tube members are controlled by companion cells (FIGURE 5-38). These cells are often smaller than the accompanying conducting cell and have a prominent nucleus and dense cytoplasm filled with ribosomes. Walls between conducting cells and controlling cells have many complex passages that are sieve areas on the conducting cell side and large plasmodesmata on the controlling cell side. Companion cells have another important role: that of loading sugars into and out of the sieve tube members.
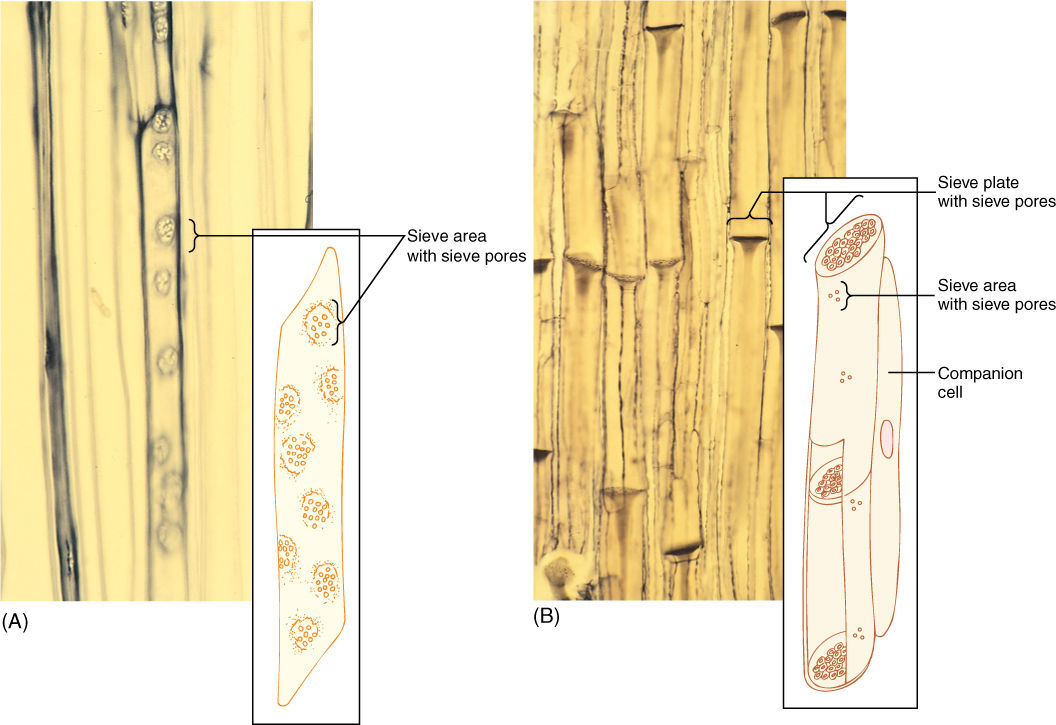
FIGURE 5-36 (A) A sieve cell is shaped like a tracheid and has sieve areas with sieve pores over much of its surface (×250). (B) A sieve tube member has flat end walls (sieve plates) with large sieve pores. On the side walls are only a few small sieve areas with very narrow sieve pores (×60). The micrograph of sieve cells is a high magnification; they are much narrower than the sieve tubes of (B).
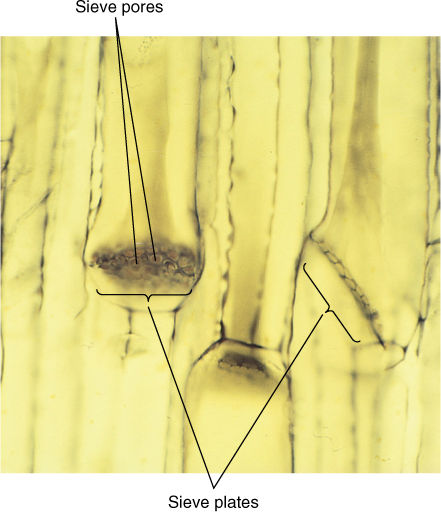
FIGURE 5-37 Large sieve areas present on sieve plates of sieve tube members. Phloem sap flows from one tube-like sieve tube member into the next through the wide sieve pores (×300).
TABLE 5-8 Sieve Elements
Vascular Bundles
Xylem and phloem occur together as vascular bundles, located just interior to the cortex (FIGURE 5-39). In basal angiosperms and eudicots, vascular bundles are arranged in one ring surrounding the pith, a region of parenchyma similar to the cortex. In monocots, vascular bundles are distributed as a complex network throughout the inner part of the stem; between the bundles are parenchyma cells. Monocot bundles are frequently described as “scattered” in the stem, suggesting that they occur at random, but they actually have precise, specific patterns that are too complex to be recognized easily.
All vascular bundles are collateral; that is, each contains both xylem and phloem strands running parallel to each other (Figure 5-39). The xylem of a vascular bundle is primary xylem because it is part of the primary plant body. In addition to conductive tracheids and vessel elements, there is usually a large proportion of xylem parenchyma and even mechanical sclerenchyma in the form of xylem fibers. The vascular bundle phloem is primary phloem, and mixed with sieve elements and companion cells or albuminous cells may be storage parenchyma and mechanical sclerenchyma, usually as phloem fibers, although phloem sclereids also occur. The relative amounts of each cell type within a bundle vary: Storage stems have large amounts of parenchyma in the vascular tissues; vines and tendrils have extra fibers.
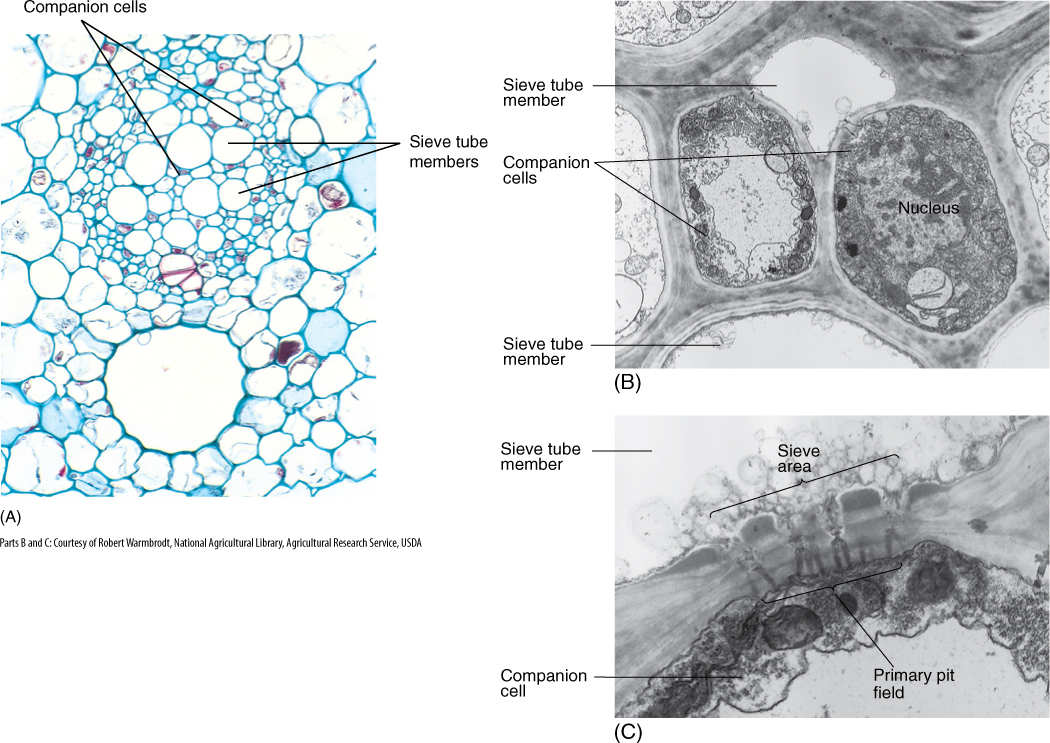
FIGURE 5-38 (A) In a vascular bundle, sieve tube members and companion cells occur together. Companion cells are small and rich in cytoplasm; sieve tube members are large and appear empty because most phloem sap is lost while preparing the tissue for microscopy (×150). (B) Electron micrograph showing two companion cells and two sieve tube members. (C) The connection between a companion cell and a sieve tube member is a primary pit field with plasmodesmata on the side of the companion cell (bottom), but it is a sieve area with sieve pores on the side of the sieve tube members (top) (×45,000).
Cells of primary xylem are larger than cells of primary phloem, and the tracheary elements on the inner side of each bundle are much smaller than the outer ones (Figure 5-39). This is true of the primary xylem in all stems, and the reason for this becomes clear when the shoot growth is examined.
![]() Stem Growth and Differentiation
Stem Growth and Differentiation
Stems grow longer by creating new cells at their tips, in regions known as shoot apical meristems; cells divide by mitosis and cytokinesis, producing progenitor cells for the rest of the stem (FIGURE 5-40). When each cell divides, the two daughter cells are half the size of the mother cell, but they grow back to the original size. As they expand, they automatically push the meristem upward; the lower cells are left behind as part of the young stem. This region just below the apical meristem is the subapical meristem, and its cells are also dividing and growing, producing cells for the region below.
In the subapical meristem, visible differentiation begins; certain cells stop dividing and start elongating and differentiating into the first tracheids or vessel elements of the vascular bundles (Figure 5-39 and FIGURE 5-41). Because they constitute the first xylem to appear, they are called protoxylem. Cells around them continue to grow and expand, but soon those just exterior to them also begin to differentiate; because these had expanded for a longer time, they develop into tracheary elements that are even larger than the first. This process continues until the last cells mature, and this portion of the stem stops elongating. Because these cells have had the longest time for growth before differentiation, they develop into the largest tracheary elements of all, called metaxylem.
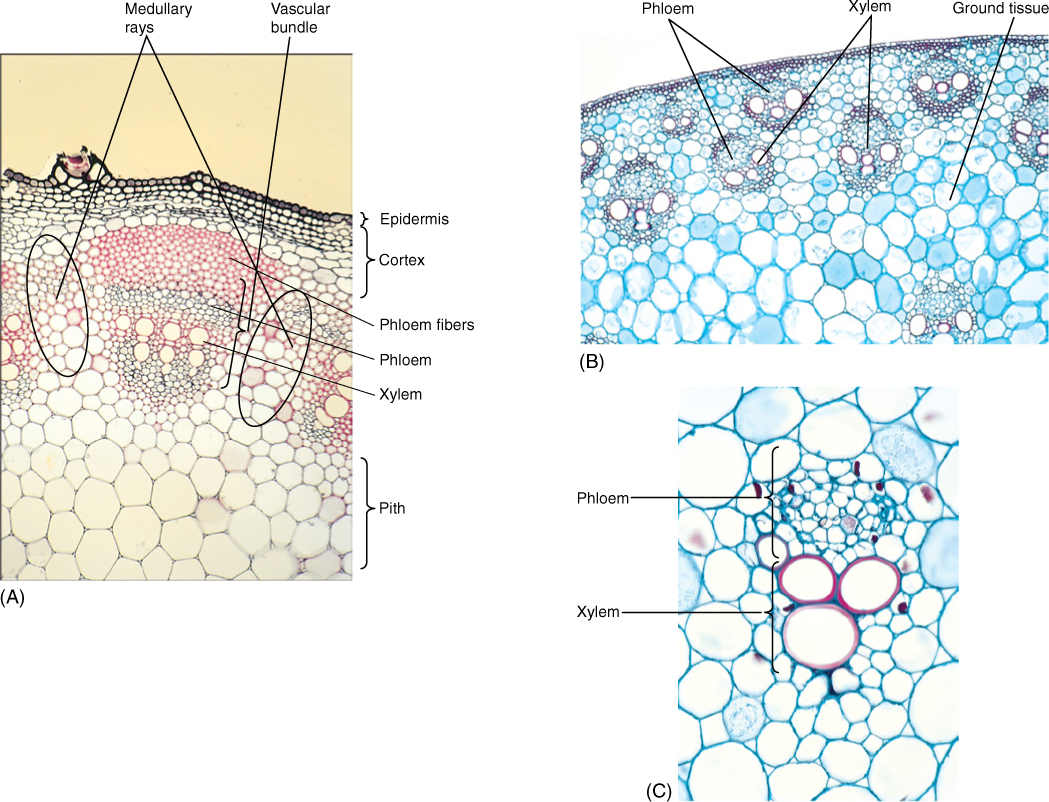
FIGURE 5-39 (A) In this vascular bundle of sunflower (Helianthus), xylem contains rows of vessels alternating with rows of xylem parenchyma. Phloem contains sieve tube members and companion cells. There is a large sclerenchyma cap of phloem fibers (×60). (B) Vascular bundles of monocots have a more complex arrangement. In this corn stem, many large bundles are located near the cortex, and fewer, smaller bundles in the ground tissue (×50). (C) High magnification of a corn bundle, showing primary xylem and phloem (×250).
Size is not the only difference between tracheary elements of protoxylem and metaxylem. Because protoxylem elements differentiate while surrounding cells are still elongating, they must be extensible and so must have either annular or helical secondary walls. Although protoxylem cells are dead at maturity, they stretch because their primary walls are glued by their middle lamellas to living, elongating cells that surround them. If protoxylem had scalariform, reticulate, or pitted secondary walls (which they never have), the cells could not be stretched; either they would prevent the surrounding cells from growing, or the middle lamellas would break and the cells would tear away from each other. This is not a problem for metaxylem, however, because its cells differentiate only after all surrounding stem tissues have stopped elongating. Because metaxylem cells do not have to be extensible, any type of secondary wall is feasible.
A similar process occurs in the outer part of each vascular bundle. The exterior cells mature as protophloem, and cells closest to the metaxylem become metaphloem. Because the sieve elements do not have secondary walls, the walls of protophloem and metaphloem are identical; however, sieve elements are extremely sensitive to being stretched and die when stressed too much. Consequently, protophloem cells are extremely short lived, often functioning for only one day. Perhaps because they are so temporary, protophloem cells never become well differentiated; their sieve areas are small and rudimentary, and many do not have companion cells. Sieve elements of metaphloem do not differentiate until later, when all surrounding cells have stopped expanding. Metaphloem cells differentiate fully, having large sieve areas and conspicuous companion cells.
You might expect metaphloem cells to be very large, like those in metaxylem, but they are much smaller, even though they have undergone long periods of expansion before differentiation. All cells of the region are expanding, whether they are dead functioning protoxylem, dead nonfunctioning protophloem, or living undifferentiated metaxylem and metaphloem. Cell size differs because cell division is still occurring in some cells but not in others. In xylem, all cells stop dividing, so each cell becomes larger as the tissue expands—protoxylem cells by being stretched and the soon-to-be metaxylem cells by their own growth. Protophloem cells stop dividing and differentiate; however, other phloem cells continue to divide, and they therefore remain small.
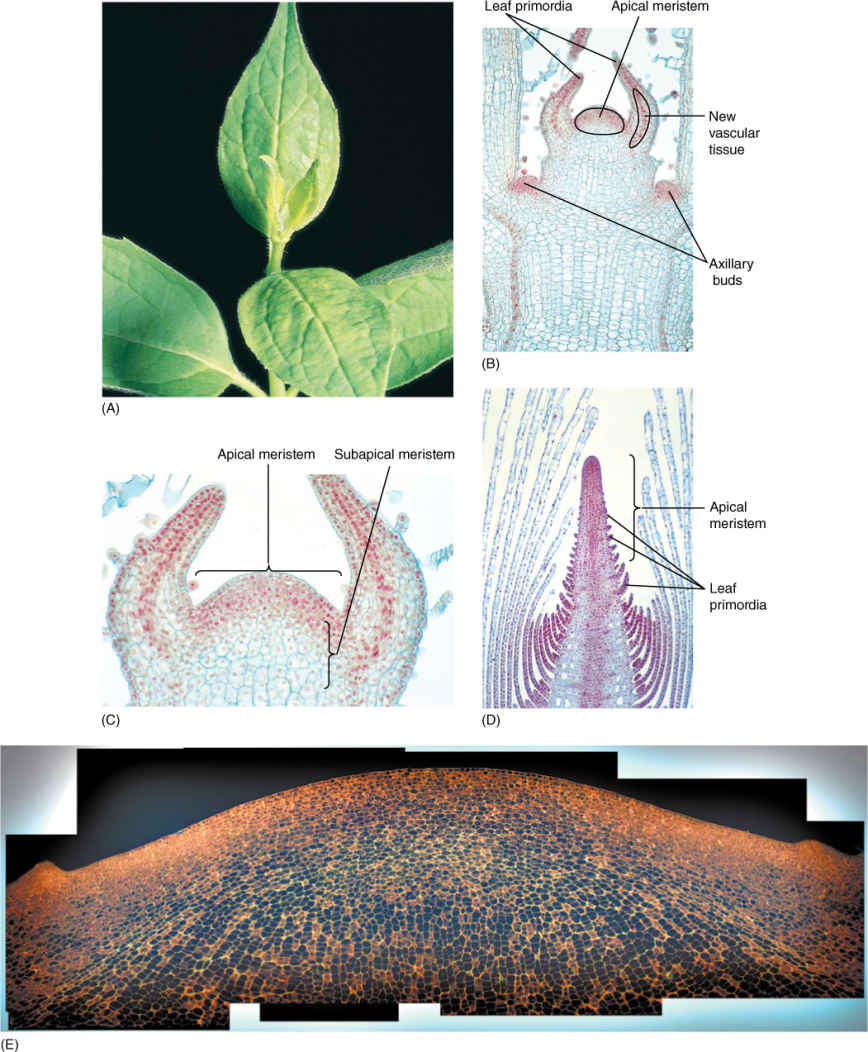
FIGURE 5-40 (A) This shoot tip has several fully expanded leaves, one set of small leaves, and another set of even smaller, younger leaves. Dissection would reveal even more, younger leaves. (B) Longitudinal section through a shoot tip of Coleus showing the apical meristem and leaf primordia of various ages. Two axillary buds are just recognizable as small outgrowths (×100). (C) Higher magnification of the Coleus shoot apical meristem. Vascular tissue is differentiating in the center of each leaf primordium (×200). (D) This apical meristem of Elodea is much taller and narrower than that of Coleus in (C), and it has many more leaf primordia (×50). (E) This is the giant shoot apical meristem of golden barrel cactus (Echinocactus grusonii); the meristem is so large it had to be photographed as a collage of approximately 15 smaller images. This image is printed at only 100 times larger than the actual meristem, which is 1.5 mm in diameter. Color was altered to enhance the visibility of cell walls and nuclei (×100).
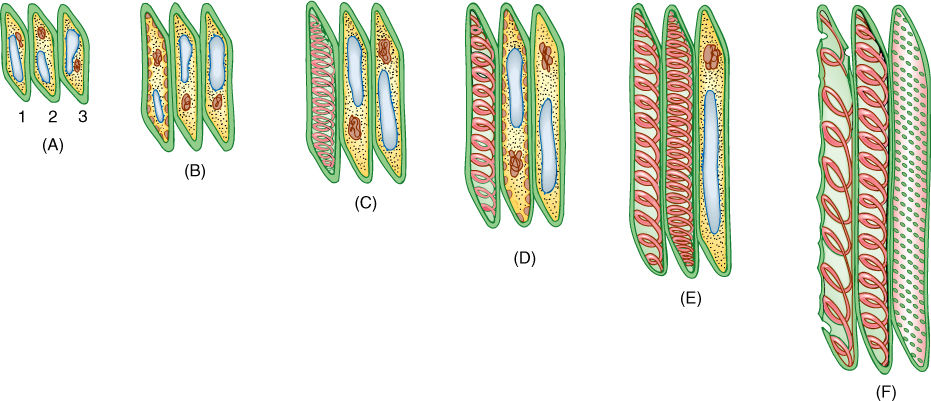
FIGURE 5-41 Development of three tracheary elements. (A) All cells are young, small, and cytoplasmic. (B) The first protoxylem element forms while all cells are still small. Notice that cell 1 has started depositing its secondary wall (red dots). (C) Cells 2 and 3 continue growing, stretching cell 1, which is dead. (D and E) Cell 2 matures while cell 3 continues to grow, stretching cells 1 and 2. Cell 1 has been stretched so much that it is on the verge of being torn open. (F) All elongation has stopped, and cell 3 differentiates as a pitted element. The primary wall of cell 1 has been torn and no longer conducts. Cell 2 is probably still capable of carrying water. In a stem, these three cells would be surrounded by numerous others.
Other tissues also differentiate in the subapical region. In the epidermis, the first stage of trichome outgrowth may be visible in the youngest internodes, those closest to the apical meristem. In lower, older internodes, trichomes are more mature, and guard cells and stomatal pores may be forming. The cuticle is extremely thin at the apical meristem, but it is thicker in the subapical region and may be complete several internodes below the shoot apex. Differentiation of the pith typically involves few obvious changes: Cells enlarge somewhat; intercellular spaces expand but remain rather small, and cell walls continue to be thin and unmodified. All changes are usually completed quickly, while the cells are in or just below the subapical meristem. Similar changes occur in the cortex, except that plastids develop into chloroplasts. Protoxylem and protophloem develop quickly while the cells are still close to the apical meristem, but metaxylem and metaphloem do not begin to differentiate until the nodes and internodes have stopped elongating.
Because the terms “epidermis,” “cortex,” “phloem,” “xylem,” and “pith” typically refer to mature cells and tissues, it is confusing to use them for the young, developing cells during differentiation. To prevent misunderstandings, a special set of terms has been agreed on: Protoderm refers to epidermal cells that are in the early stages of differentiation. Young cells of xylem and phloem are provascular tissues; the equivalent stages of pith and cortex are ground meristem (FIGURE 5-42). Thus, the subapical meristem consists of protoderm, ground meristem, provascular tissue, and more ground meristem. The rate of maturation varies greatly from species to species and also from one type of stem to another on the same plant. In stems with long internodes, such as vines, maturation is slow, and immature tissues can be found far below the shoot apical meristem. In stems with short internodes, such as bulbs and lettuce, differentiation may be completed quickly, and all tissues are mature close to the stem apex.
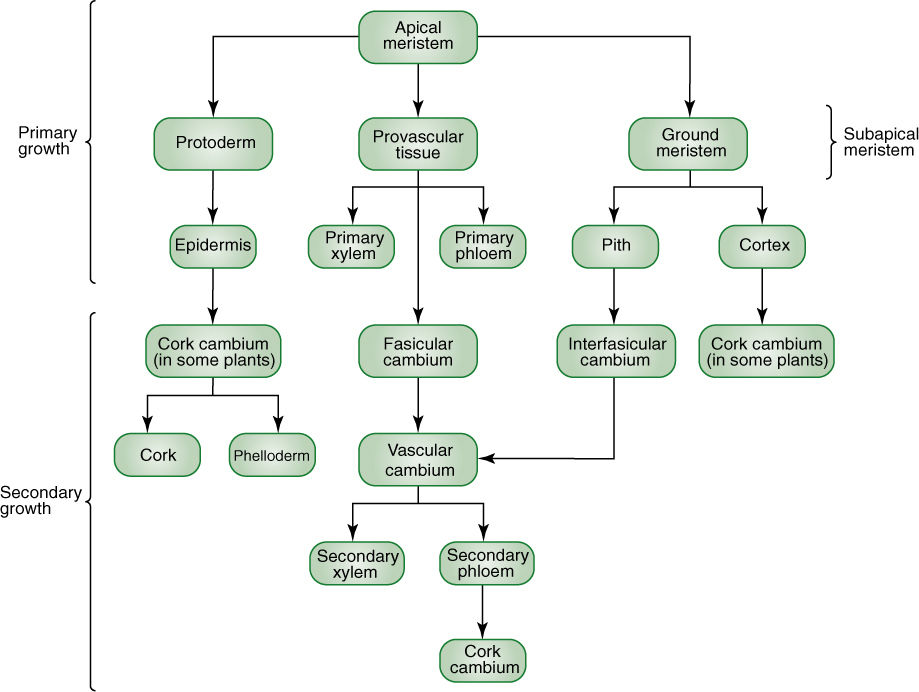
FIGURE 5-42 Cells of an apical meristem give rise to three types of subapical cells, which in turn give rise to particular types of mature primary tissues. Epidermis develops only from protoderm, never from provascular tissue or ground meristem; other cell lineages in the primary body are also quite simple and linear.
Now, at the end of the chapter, we can define two concepts precisely. Primary tissues are the tissues produced by apical meristems; primary growth is the growth and tissue formation that results from apical meristem activity. Secondary tissues (wood and bark) are produced by different type of meristems (vascular and cork cambia) and are discussed elsewhere.
Plants Do Things Differently
BOX 5-4 Plants and People Grow Differently
Imagine that your parents planted a tree seed on the day you were born. That tree is probably still getting taller while your growth in height has come to an end. We achieve our maximum height when we are approximately 18 years old, but trees keep right on growing. The tips of their twigs lengthen several inches every spring and make new leaves and flowers. How tall will the tree become? If we treat them well, give them fertilizer, water, sunlight, and protect them from diseases and storms, they may never stop growing. For us, however, our height is set. We will not get to be 7 or 8 feet tall no matter how many vitamins we take, no matter how nutritious our food.
We animals have determinate growth. Plants have indeterminate growth. The maximum amount of growing an animal will do in its lifetime, and therefore, its size and the number of organs it has are determined by the genes inherited from its parents. Malnourishment may prevent us from growing properly, but even if healthy and well nourished, we cannot exceed our genetic potential. The size of most plants is not determined by their genes because even if a plant received genes for poor growth, it might still become extremely large if it lives an unusually long time.
An animal’s determinate growth is related to its body’s precise, invariant form with a specific number of organs. We have one head, one torso, two arms, two legs, and our spine contains 33 vertebrae and 12 pairs of ribs, and these were all present when we were born. No new ones were added later: We have determinate organogenesis. Plant bodies are not so precisely organized. When a plant germinates as a seedling, most of its leaves and roots are not present yet, and it has none of its branches, flowers, or fruits either. All of these will be made over the many years of its life, and there is no set number of organs. There will never be a time when it has all of its leaves because some will have already fallen off. Others will not have been made yet. Plants have indeterminate organogenesis.
Plants grow by special groups of cells at the tips of their shoots and roots. As these cells divide, approximately half the new cells mature and differentiate into functional cells of the stem, new leaves, and flowers and so on; however, the other cells simply grow back to their original size then divide again. Once again, about half the new cells mature, and the other half continue as cells capable of dividing. The shoot tip always has cells able to generate new cells. There is no reason for it to ever run out of cells. Animals have no set of cells like this, cells always capable of making new cells and organs for the body’s growth.
Growth is not one hundred percent different in plants and animals, however. Our skin’s growth is indeterminate. The innermost layer of our skin, called the stratum basale, consists of cells that never stop dividing; after each round of division, one daughter cell matures as a skin cell, and the other continues to divide. We replace our skin continuously. After a bad sunburn, some skin peels off, but the stratum basale makes more. This is the way plants grow. Our hair, fingernails, and blood also grow continuously from sets of cells that divide and provide new cells throughout our life. Becoming more muscular or fatter is different. That just involves making existing cells larger, not increasing their numbers.
Leaf growth in plants is determinate. While a leaf is small, all its cells divide repeatedly. Then, at some point, all differentiate and mature, and the leaf has no cells capable of making any new leaf cells. It expands to its full size as each cell enlarges its central vacuole. The leaf’s maximum size is determined by the plant’s genes. The same is true for flowers. If we give our plants good care, they may make more flowers but not larger ones.
Now for the exceptions: Tapeworms have indeterminate growth. They have a “head” that attaches to the host’s intestine and a “neck” that is a set of cells that divide repeatedly. When enough new cells are present, some organize themselves into a body segment complete with organs. The dividing neck cells continue to make more cells for another segment. The tapeworm makes an indeterminate number of segments, growing to an indeterminate length. Older segments full of eggs break off and end up in the host’s excrement, but the “neck” makes new replacement segments. Unlike other animals, tapeworms do not have a specific number of parts other than one head and one neck. Its plant-like indeterminate growth is based on its plant-like set of permanently dividing cells and plant-like lack of precise body pattern.
Exceptional plants are annual plants. These live for just 1 year and, like animals, have determinate growth. Even though they grow at their tip, for some reason those cells all stop dividing after a few weeks or months, ending the plant’s organ formation. The plant then blooms and makes fruits and seeds, then it dies. Biennial plants live for 2 years, making many leaves in the first year, blooming and reproducing in the second and then dying. The life of annuals and biennials is programmed by their genes. No matter how much care and fertilizer we give them, we cannot keep them alive longer or get them to grow indeterminately.
 At the Next Level
At the Next Level
1. Extreme modification of shoots. All seed plants have the same basic organization of their shoots, but in some, the various tissues are highly modified. Although cacti appear to be exceptional, they have all the normal parts presented in this chapter; you now know enough to understand papers about their anatomy. Parasitic plants that live entirely within their hosts usually have such highly modified bodies that some organs and tissues are missing or barely recognizable. Species you might find informative are Rafflesia arnoldii, Tristerix aphyllus, Viscum minimum, and Hydnora africana. If you really want to go to the next level, look up the family Podostemaceae (sometimes named Podostemonaceae): These are flowering plants in which almost all aspects of anatomy are abnormal.
2. Evolution and ecology of xylem. All organisms must obtain, transport, and conserve water. Many studies have analyzed the ways in which alternative types of xylem cells affect a plant’s adaptation to its habitat. Also the evolution of xylem—from a few simple tracheids in the first plants to the many diverse types of cells present in angiosperm xylem—has been studied extensively and is well-documented. Searching on the key words “xylem evolution” and “xylem ecology” is a good way to start.
3. Ant plants (myrmecophytes). A surprising number of plants have stem modifications, such as hollow chambers with tunnels leading to the shoot surface, that provide shelter for animals such as ants and mites. The structures are called domatia (sing.: domatium; pronounced doe MAY shum) and the insects that occupy them typically provide some sort of benefit to the plant. For example ants often patrol the plant and kill any other insects that would attack the plant, and some even clean fungi off the plant. Many also urinate and defecate in special chambers of the domatia, thus providing the plant with nitrogen fertilizer. Search on these keywords: ant plant, myrmecophyte, domatium, Hydnophytum, Myrmecodia, Solanopteris (and check the index of this book).
SUMMARY
1. Cells must be arranged in the proper patterns and with the proper interrelationships in order to function efficiently.
2. All angiosperms have roots, stems, and leaves, although each can be highly modified in particular environments.
3. Virtually all stems function in producing and supporting leaves and transporting water, minerals, and sugars between leaves, buds, and roots. Other stems are involved in storage, reproduction, dissemination, and surviving stress.
4. All stems consist of nodes and internodes, and they have leaves and axillary buds. Numerous types of stems exist; most individual plants have two or three types of shoots.
5. The three basic types of plant cells, based on walls, are parenchyma (responsible for most metabolism), collenchyma (plastic support), and sclerenchyma (elastic support).
6. The vascular tissues are xylem and phloem. Tracheary elements consist of vessel elements (with perforations) and tracheids (without perforations). Sieve elements consist of sieve cells (with small sieve pores) and sieve tube members (with large sieve pores on the end walls).
7. Stems of all angiosperms have epidermis, cortex, and collateral vascular bundles. In basal angiosperms and eudicots, the bundles occur as one ring surrounding pith, whereas in monocots, they have a complex distribution in conjunctive tissue.
8. Protoxylem and protophloem form while an organ is elongating and therefore must be extensible. Metaxylem and metaphloem form after elongation ceases.
9. Both primary xylem and primary phloem are complex tissues with a variety of cell types, not just conducting cells.
IMPORTANT TERMS
angiosperms
apical meristem
axillary bud
basal angiosperms
bulbs
circular bordered pits
collenchyma cells
companion cells
corms
cortex
cuticle
epidermis
eudicots
fibers
ground meristem
guard cells
internodes
metaphloem
metaxylem
monocots
nodes
parenchyma
perforation
phloem
phyllotaxy
pits
pit membrane
pit-pair
pith
primary growth
primary tissues
protoderm
protophloem
protoxylem
provascular tissues
rhizomes
sclereids
sclerenchyma
sieve areas
sieve cells
sieve plates
sieve pores
sieve tube
sieve tube members
stoma
tracheary element
tracheids
trichomes
tubers
vascular bundles
vessel
vessel elements
xylem
REVIEW QUESTIONS
1. The body of an herb contains just three basic parts. What are they?
2. Stems of the first land plants functioned primarily as transport and support structures. Modern stems have these functions also, plus several more. Describe the additional functions.
3. Over the years, several different names have been used for the flowering plants. Here they are called the division_________________________, but they are known informally as _________________________. There are about __________________ species of flowering plants.
4. Early angiosperms diversified into several groups that are now known as,____________ _____________, and _________________________. Name three examples of plants within each line of evolution.
5. An herb has only a primary plant body. That means it has roots, stems, and leaves, but it never becomes _________________________ and covered with _________________________.
6. In general, animals have _________________________ growth with a fixed size and number of organs, whereas plants have _________________________ growth with no set size and any number of organs. Which parts of our body grow continuously like plants? Which parts of a plant have fixed size such that no matter how well we take care of the plants those organs do not become larger? What is the name of plants that live for just 1 year? For just 2 years?
7. What are the important differences between parenchyma, collenchyma, and sclerenchyma cells?
8. Examine Figure 5-6A and 5-6B. Why do most parenchyma cells shown in Figure 5-6A lack nuclei, whereas most shown in Figure 5-6B do have nuclei? In the geranium for Figure 5-6A, did the real cells lack nuclei, or did something happen to the nuclei as the tissue was sliced to make the microscope slide?
9. What is the special name of photosynthetic parenchyma cells? Which organelle is especially abundant in these? How does wall thinness affect carbon dioxide and light?
10. It was previously stated that parenchyma cells are relatively inexpensive to build. What does that mean? How does this relate to most leaves being soft?
11. Like clay, walls of collenchyma exhibits plasticity. Does that mean it can or cannot be stretched? If a tissue is supported by collenchyma, can it still grow?
12. How does collenchyma support a tissue? Would it work if the tissue did not have parenchyma also? What happens if the tissue is not turgid (does not have enough water)?
13. Collenchyma is plastic, but sclerenchyma is elastic. What does “elastic” mean? If you stretch or deform an elastic object, will it keep its new shape or snap back to its original shape? As an organ grows, its shape changes, but after it has achieved its mature size and shape, what kind of things would deform it? Should a mature organ have plastic properties or elastic ones (e.g., if a heavy load of snow bends a branch down, should the branch stay in its bent shape when the snow melts or should it go back to the shape it grew to before the snow)?
14. What are the two types of mechanical, nonconducting sclerenchyma (hint: Table 5-3)? Which tends to be flexible and useful in wood? Which tends to be brittle and inflexible, useful in “pits” and “stones” that protect seeds?
15. Fibers and sclereids have secondary walls that are so thick and tough that the cell cannot grow. If that is true, how do fibers and sclereids grow to their mature size and shape? Do they have a secondary wall even when they are young?
16. Imagine a leaf of a palm tree. When it is fully grown, it must have sclerenchyma to support its size and weight. Do you think it has any sclerenchyma when it is tiny and just starting to grow? Would you expect that it might have collenchyma when it is medium sized and still growing?
17. Technically, the _________________________ is an axis, whereas the _________________________ is the _________________________ plus any leaves, flowers, or buds that might be present.
18. The point where a leaf is attached to a stem is called a _________________________. Just above this point is an _________________________ bud.
19. Figure 5-13A shows two types of buds. What are the two types and how do they differ?
20. After a leaf falls off a stem, it leaves a _________________________ just below the axillary bud.
21. What is phyllotaxy? Corn and irises have two rows of leaves. This is known as _________________________ phyllotaxy.
22. When you look at a head of cabbage or lettuce, what are you seeing? Where is the stem? What about when you look at an onion? What are you seeing and where is the onion’s stem?
23. Describe each of the following types of specialized shoots. Be certain to account for modifications of the leaves, internodes, and orientation of growth: stolon, rhizome, tuber, bulb, corm, and tendril. Each provides a plant with a selective advantage. What is the adaptive value of each type of specialized shoot?
24. Many scientific words are also ordinary English words. If you read that a plant has a bulb or a tuber, can you always be certain exactly what the botanical structure is?
25. What is the outermost surface of an herbaceous stem? The outer walls of this layer are encrusted with a chemical made up of fatty substance that makes the wall impermeable to water. What is the name of the substance and what is the name of the layer?
26. A stoma allows carbon dioxide to pass through the epidermis. What is the name of the two cells that control the opening and closing of a stoma? What is the name of the hole itself that carbon dioxide passes through?
27. What is the technical term for a plant hair? Plant hairs make it difficult for animals to do certain things. Name three activities that are more difficult for an animal because of a hairy leaf.
28. What is the name of the region of cells between a stem’s epidermis and its set of vascular bundles? This is usually a compact parenchyma tissue, but in some aquatic angiosperms that live submerged in lakes or oceans, this region has what type of special modification? Would you hypothesize that this region is especially thick or thin in desert plants such as cacti or plants that have succulent water storage tissues (you must think about this yourself, as the answer is not in the text).
29. All flowering plants have two types of vascular tissues, _________________________, which conducts water and minerals, and _________________________ , which distributes sugars and minerals. Are vascular systems of plants similar to or different from those of animals? Explain the similarity or differences.
30. As a young xylem cell matures into a tracheary element, it first must enter _________________________ and stop _________________________. It is initially a small _________________________ cell, but after it reaches its full size and shape, it deposits a _________________________ wall that reinforces the wall. The xylem cell _________________________ and its protoplasm _________________________, leaving a _________________________ wall.
31. List the five types of secondary wall deposition that can occur in tracheary elements. Which two types are most characteristic of protoxylem? What is the selective advantage of vessel elements over tracheids?
32. Consider tracheary elements with annular thickening (annular secondary walls) and those with pitted walls. Which is weaker? Which has a large percentage of its primary wall free of secondary wall and available for water movement? Is one always more selectively advantageous than the other? Under all conditions? Explain.
33. Tracheids obtain water from _________________________ below them and pass it on to _________________________ above. As water passes into or out of a tracheid, it passes through a _________________________ - _________________________. The set of primary walls and middle lamella in the _________________________ - _________________________ is called a _________________________.
34. If each tracheid is 1 mm long, how many pit-pairs and pit membranes will a water molecule pass through in a plant 1 m tall? Is this a significant amount of friction?
35. What is a perforation in a vessel element? How does it reduce friction as water passes from one vessel element to another? When a water molecule arrives at the end of a vessel, how does it pass into another vessel (this is asking about vessels, not vessel elements). In Question 34, tracheids were assumed to be 1 mm long. If we assume that vessel elements are also 1 mm long (they are usually shorter, but 1 mm is an easy number to work with), how many vessel elements are there in an average vessel of American beech (Table 5-7, the lengths in the table are in centimeters)? The plant in Question 34 was said to be 1 m tall. Would an average vessel of American beech even fit into a plant this tall? If a plant 1 m tall had vessels 1 m long, how many pit membranes would a water molecule need to pass through as it traveled from root to shoot tip? Is this more or less than the answer for Question 34?
36. Which evolved more recently, tracheids or vessel elements? Do nonangiosperms such as conifers and ferns have vessel elements? Do flowering plants (angiosperms) have tracheids? Vessel elements?
37. Like xylem, phloem has two types of conducting cells, _________________________ and _________________________. The term “_________________________ _________________________” refers to either one. Do these die like tracheary elements or do they need to remain alive in order to conduct?
38. What is the name of the holes that interconnect conducting cells in phloem, and what are groups of these holes called?
39. Sieve tube members differ from sieve cells by having very large sieve pores on their end walls, much larger than the ones on their side walls. What is the name of these end-wall sieve areas with big sieve pores?
40. Sieve elements (both sieve cells and sieve tube members) lose their nuclei during development, but they must remain alive. What is the name of the cell associated with sieve cells? The one associated with sieve tube members?
41. During differentiation of a young cell into a sieve tube member, what are some changes that occur in the cell wall and the cytoplasm? How long do most sieve tube members live after they become mature?
42. Describe plasmodesmata, pits, perforations, and sieve pores. Which connect living cells? Which connect nonliving cells? Which occur in secondary walls?
43. All vascular bundles are _________________________. That is, each contains both xylem and phloem. The xylem of a vascular bundle is _________________________ xylem because it is part of the primary plant body, and the phloem is _________________________ phloem.
44. Stems grow longer by creating new cells at their tips, in regions known as _________________________. Below this region, in the subapical meristem, the very first primary xylem to appear is called _________________________, and cells that differentiate into xylem a little later, after they have grown larger, are called _________________________.
45. In the apical and subapical region, there are cells that will later give rise to epidermis and other tissues, but they are not yet mature enough to call them epidermis, and so on. What are the terms for the following cells?
a. Epidermal cells that are still meristematic
b. Young cells of xylem and phloem
c. Young cells of pith and cortex
46. Describe the arrangement of tissues seen in a stem cross-section; consider monocots separately from the other angiosperms. Is the arrangement different in a stolon than in a rhizome, tuber, or corm?
47. Look at Figure 5-42. The top three rows are labeled “Primary growth” because these are found in the primary plant body, the body of an herb. The third row of the figure has five boxes with the five types of mature tissue found in the primary plant body. What are the five tissues?
48. If you live to be 100 years old, how many sets of organs will you have during your lifetime? How many hearts, stomachs, and livers? If a plant lives to be 100 years old, how many sets of leaves will it probably have? If it begins blooming when it is 10 years old, how many sets of flowers? When you are 70 years old, you will still depend on your digestive system to supply you with energy for another 30 years, but how old is that set of organs? In a 70-year-old tree, how old are the cells of the leaves that supply it with energy?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields;
Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art