Biology For Dummies
Part II Cell Reproduction and Genetics: Let’s Talk about Sex, Baby

In this part . . .
Living things grow and reproduce both sexually (through meiosis) and asexually (through mitosis). They also tend to pass their traits on to their offspring. The blueprints for these traits are encoded in the structure of the most famous molecule of life: deoxyribonucleic acid, or DNA. Scientists today know more about the mysteries of DNA than ever before. They’ve even figured out ways to harness its power for medicine, agriculture, and technology.
In this part, we explain the steps of cell division and how DNA determines the characteristics of organisms.
Chapter 6
Dividing to Conquer: Cell Division
In This Chapter
Understanding why cells reproduce and how DNA replicates itself
Discovering how mitosis produces exact copies of cells
Making egg and sperm cells through meiosis
Appreciating the power of genetic diversity
All living things can reproduce their cells for growth, repair, and reproduction. Asexual reproduction by mitosis creates cells that are genetically identical to the parent cell. Sexual reproduction requires a special type of cell division called meiosis that produces cells containing half the genetic information of the parent cell. Meiosis and sexual reproduction result in greater genetic diversity in offspring and, consequently, in the populations of living things. In this chapter, we explore the reasons why cells divide and present the steps of each type of cell division. We also introduce you to the ways sexual reproduction adds variety to species of all kinds.
Reproduction: Keep On Keepin’ On
Biology is, of course, all about life. And, when you think about it, life is really all about continuation — living things keep on keepin’ on from one generation to the next, passing on critical genetic information. Certainly this is one of the core differences between organisms and inanimate objects. After all, have you ever seen a chair or table replicate itself? Only living things have the ability to pass on genetic information and replicate themselves.
When cells replicate, they make copies of all of their parts, including their DNA, and then divide themselves to make new cells. If a cell makes an exact copy of itself, it’s engaging in asexual reproduction. Single-celled prokaryotes, such as bacteria, reproduce asexually by binary fission;they’re able to divide quickly and reproduce themselves in as little as 10 to 20 minutes. Some single-celled eukaryotes and individual cells within a multicellular eukaryote also reproduce asexually. However, they use a process called mitosis (which we explain in the later “Mitosis: One for you, and one for you” section) to produce new generations. If a cell produces a new cell that contains only half of its genetic information, that cell has engaged in sexual reproduction. A special type of cell division known as meiosis (which we explain in the later “Meiosis: It’s all about sex, baby” section) is responsible for all sexual reproduction.
 Cells divide for the following important reasons:
Cells divide for the following important reasons:
To make copies of cells for growth: You started out as a single cell after mom’s egg met dad’s sperm, but today you have about 10 trillion cells in your body. All of those cells were produced from that first cell and its descendents by mitosis. When you watch plants grow taller or baby animals grow into adults, you’re seeing mitosis at work.
To make copies of cells for repair: It’s a fact of life that cells wear out and need to be replaced. For instance, you constantly shed skin cells from your surface. If your body couldn’t replace these cells, you’d run out of skin. And if an organism gets injured, its body uses mitosis to produce the cells necessary to repair the injury.
To carry on the species: During asexual reproduction, organisms make exact copies of themselves for the purpose of creating offspring. During sexual reproduction, gametes (cells, specifically eggs and sperm, containing half the genetic information of their parent cells) get together to make new individuals. When the genetic information of the gametes joins together, the new individual has the correct total amount of DNA.
Welcome to DNA Replication 101
If one cell is going to divide to produce two new cells, the first cell must copy all of its parts before it can split in half. The cell grows, makes more organelles (see Chapter 4 for the full scoop on organelles), and copies its genetic information (the DNA) so that the new cells each have a copy of everything they need. Cells use a process called DNA replication to copy their genetic material. In this process, the original DNA strands serve as the template for the construction of the new strands. It’s particularly important that each new cell receives an accurate copy of the genetic information because this copy, whether it’s accurate or faulty, directs the structure and function of the new cells.
The basic steps of DNA replication are as follows:
First, the two parental DNA strands separate so that the rungs of the double helix ladder are split apart with one nucleotide on one side and one nucleotide on the other (see Chapter 3 for a depiction of a DNA molecule). The entire DNA strand doesn’t unzip all at once, however. Only part of the original DNA strand opens up at one time. The partly open/partly closed area where the replication is actively happening is called the replication fork (this is the Y-shaped area in Figure 6-1).
The enzyme DNA polymerase reads the DNA code on the parental strands and builds new partner strands that are complementary to the original strands. To build complementary strands, DNA polymerase follows the base-pairing rules for the DNA nucleotides: A always pairs with T, and C always pairs with G (see Chapter 3 for further details about nucleotides). If, for example, the parental strand has an A at a particular location, DNA polymerase puts a T in the new strand of complementary DNA it’s building. When DNA polymerase is done creating complementary pairs, each parental strand has a brand-new partner strand.
DNA polymerase is considered semiconservative because each new DNA molecule is half old (the parental strand) and half new (the complementary strand).
Several enzymes help DNA polymerase with the process of DNA replication (you can see them and DNA polymerase hard at work in Figure 6-1):
Helicase separates the original parental strands to open the DNA.
Primase puts down short pieces of RNA, called primers, that are complementary to the parental DNA. DNA polymerase needs these primers in order to get started copying the DNA.
DNA polymerase I removes the RNA primers and replaces them with DNA, so it’s slightly different from the DNA polymerase that makes most of the new DNA (that enzyme is officially called DNA polymerase III, but we refer to it simply as DNA polymerase).
DNA ligase forms covalent bonds in the backbone of the new DNA molecules to seal up the small breaks created by the starting and stopping of new strands.
The parental strands of the double helix are oriented to each other in opposite polarity: Chemically, the ends of each strand of DNA are different from each other, and the two strands of the double helix are flipped upside down relative to one another. Note in Figure 6-1 the numbers 5' and 3' (read “5 prime” and “3 prime”). These numbers indicate the chemical differences of the two ends. You can see that the 5' end of one strand lines up with the 3' end of the other strand. The two strands of DNA have to be flipped relative to each other in order for the bases that make upthe rungs of the ladder to fit together the right way for hydrogen bonds to form between them. Because the two strands have opposite polarity, they’re antiparallel strands.
Figure 6-1:DNA replication.
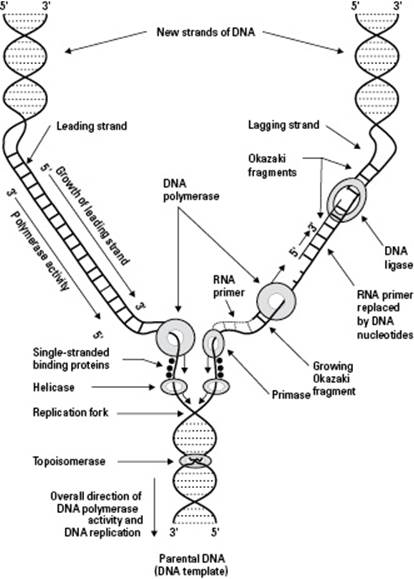
The antiparallel strands of the parent DNA create some problems for DNA polymerase. One quirk of DNA polymerase is that it’s a one-way enzyme — it can only make new strands of DNA by lining up the nucleotides a certain way. But DNA polymerase needs to use the parent DNA strands as a pattern, and they’re going in opposite directions. As a result, DNA polymerase makes the two new strands of DNA a bit differently from each other, as you can see from the following:
One new strand of DNA, called the leading strand, grows in a continuous piece. Refer to Figure 6-1. See how the new DNA on the left side of the replication fork is growing smoothly? The 3' end of this new strand points toward the replication fork, so after DNA polymerase starts building the new strand, it can just keep going.
One new strand of DNA, called the lagging strand, grows in fragments. Look at Figure 6-1 again. Notice how the right side of the replication fork looks a little messier? That’s because the replication process doesn’t occur smoothly over here. The 3' end of this new strand points away from the fork. DNA polymerase starts making a piece of this new strand but has to move away from the fork to do so (because it can only work in one direction). DNA polymerase can’t go too far from the rest of the enzymes that are working at the fork, however, so it has to keep backing up toward the fork and starting over. As a result, the lagging strand is made in lots of little pieces called Okazaki fragments. After DNA polymerase is done making the fragments, the enzyme DNA ligase comes along and forms covalent bonds between all the pieces to make one continuous new strand of complementary DNA.
Cell Division: Out with the Old, In with the New
Cell division is the process by which new cells are formed to replace dead ones, repair damaged tissue, or allow organisms to grow and reproduce. Cells that can divide spend some of their time functioning and some of their time dividing. This alternation between not dividing and dividing is known as the cell cycle, and it has specific parts:
The nondividing part of the cell cycle is called interphase. During interphase, cells are going about their regular business. If the cell is a single-celled organism, it’s probably busy finding food and growing. If the cell is part of a multicellular organism, like a human, it’s busy doing its job. Maybe it’s a skin cell protecting you from bacteria or a fat cell storing energy for later.
Cells that receive a signal to divide enter a division process, which is either mitosis or meiosis.
• Cells that reproduce asexually, like a skin cell that needs to replace some of your lost skin, divide by mitosis, a process that produces cells that are identical to the parent cell.
• Cells that reproduce sexually enter a special process called meiosis that produces special cells called gametes (in animals) and spores (in plants, fungi, and protists) that have half the genetic information of the parent cell. In you, the only cells that reproduce by meiosis are cells in your gonads. Depending on your gender, your gonads are your testes or your ovaries. Cells in testes produce gametes called sperm, and cells in ovaries produce gametes called eggs.
Mitosis and meiosis have many similarities, but the differences are essential. We cover both processes (as well as the interphase) in the sections that follow, but Table 6-1 can help you sort out the important differences at a glance.
|
Table 6-1 A Comparison of Mitosis & Meiosis |
|
|
Mitosis |
Meiosis |
|
One division is all that’s necessary to complete the process. |
Two separate divisions are necessary to complete the process. |
|
Chromosomes don’t get together in pairs. |
Homologous chromosomes must synapse to complete the process, which occurs in prophase I. |
|
Homologous chromosomes don’t cross over. |
Crossing-over is an important part of meiosis and one that leads to genetic variation. |
|
Sister chromatids separate in anaphase. |
Sister chromatids separate only in anaphase II, not anaphase I. (Homologous chromosomes separate in anaphase I.) |
|
Daughter cells have the same number of chromosomes as their parent cells, meaning they’re diploid. |
Daughter cells have half the number of chromosomes as their parent cells, meaning they’re haploid. |
|
Daughter cells have genetic information that’s identical to that of their parent cells. |
Daughter cells are genetically different from their parent cells. |
|
The function of mitosis is asexual reproduction in some organisms. In many organisms, mitosis functions as a means of growth, replacement of dead cells, and damage repair. |
Meiosis creates gametes or spores, the first step in the reproductive process for sexually reproducing organisms, including plants and animals. |
Interphase: Getting organized
During interphase, cells engage in the metabolic functions that make them unique. For instance, nerve cells send signals, glandular cells secrete hormones, and muscle cells contract. If cells get a signal to reproduce themselves, they grow, copy all of their structures and molecules, and make the structures they need to help cell division proceed in an organized fashion (inter- means “between,” so interphase is literally the phase between cell divisions).
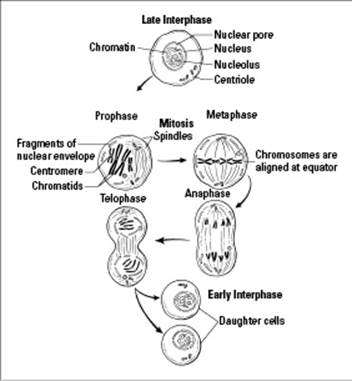
The nuclear membrane is intact throughout interphase, as you can see in Figure 6-2. The DNA is loosely spread out, and you can’t see individual chromosomes. Cells that are going to divide copy their DNA during interphase.
Interphase has three subphases:
G1 phase: During this phase, which is typically the longest one of the entire cell cycle, the cell grows and produces cell components. Each chromosome is made up of just a single double-stranded piece of DNA (double-stranded is just another way of saying that the DNA is a double helix).
Some cells actually never leave the G1 phase. They never divide; instead, they just hang out and do their cellular thing. Human nerve cells are perfect examples of cells that never leave the G1 phase.
S phase: This phase is when the cell gets ready to divide and puts the pedal to the metal for DNA replication. Every DNA molecule is copied exactly, forming two sister chromatids (a pair of identical DNA molecules) that stay attached to each other in each replicated chromosome. You can see replicated chromosomes in Figure 6-2, in the cell labeled Prophase. Each replicated chromosome looks like an X, and each X represents two identical sister chromatids held together at a location on the chromosome called the centromere.
G2 phase: During this phase, the cell is packing its bags and getting ready to hit the road for cell division by making the cytoskeletal proteins it needs to move the chromosomes around. When you look at cells that are dividing, the cytoskeletal proteins look like thin threads, hence their name — spindle fibers. A network of spindle fibers spreads throughout the cell during mitosis to form the mitotic spindle, which is represented by the thin curving lines drawn in the cells in Figure 6-2. The mitotic spindle organizes and sorts the chromosomes during mitosis.
Figure 6-2:Interphase and mitosis.
Mitosis: One for you, and one for you
After interphase is over (see the preceding section), cells that are going to divide to create an exact replica of a parent cell enter mitosis. During mitosis, the cell makes final preparations for its impending split. Processes during mitosis ensure that genetic material is distributed equally so each new daughter cell receives identical information (eukaryotic cells are model parents intent on avoiding bickering between their daughter cells).
The process of mitosis occurs in four phases, with the fourth phase initiating a final process called cytokinesis. We outline everything for you in the following sections.
Examining the four phases of mitosis
Although the cell cycle is a continuous process, with one stage flowing into another, scientists divide the events of mitosis into four phases based on the major events in each stage. The four phases of mitosis are
Prophase: The chromosomes of the cell get ready to be moved around by coiling themselves up into tight little packages. (During interphase, the DNA is spread throughout the nucleus of the cell in long thin strands that would be pretty hard to sort out.) As the chromosomes coil up, or condense, they become visible to the eye when viewed through a microscope. During prophase:
• The chromosomes coil up and become visible.
• The nuclear membrane breaks down.
• The mitotic spindle forms and attaches to the chromosomes.
• The nucleoli break down and become invisible.
Metaphase: The chromosomes are tugged by the mitotic spindle until they’re all lined up in the middle of the cell. (Meta- means “middle,” so it’s officially metaphase when the chromosomes are lined up in the middle; see the cell labeled Metaphase in Figure 6-2.)
Anaphase: The replicated chromosomes separate so that the two sister chromatids (identical halves) from each replicated chromosome go to opposite sides (see the cell labeled Anaphase in Figure 6-2). This way each new cell has one copy of each DNA molecule from the parent cell when cell division is over.
Telophase: The cell gets ready to divide into two by forming new nuclear membranes around the separated sets of chromosomes. The two daughter nuclei each have a copy of every chromosome that was in the parent cell, as you can see in Figure 6-2.
 The events of telophase are essentially the reverse of prophase.
The events of telophase are essentially the reverse of prophase.
• New nuclear membranes form around the two sets of chromosomes.
• The chromosomes uncoil and spread throughout the nucleus.
• The mitotic spindle breaks down.
• The nucleoli reform and become visible again.
Seeing how daughter nuceli go their own way with cytokinesis
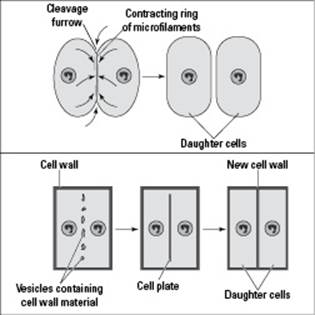
The last order of cell-division business is to give the new daughter nuclei their own cells through a process called cytokinesis. (Cyto- means “cell,” and kinesis means “movement,” so cytokinesis literally means “moving cells.”) Cytokinesis occurs differently in animal and plant cells, as you can see from the following list and Figure 6-3:
In animal cells, cytokinesis begins with an indentation, called a cleavage furrow, in the center of the cell. Cytoskeletal proteins act like a belt around the cell, contracting down and squeezing the cell in two (imagine squeezing a ball of dough at the center until it becomes two balls of dough).
In plant cells, a new cell wall forms at the center of the cell. Because a rigid cell wall is involved, the cell can’t be squeezed in two. Instead, vesicles deliver wall material to the center of the cell and then fuse together to form the cell plate. The vesicles are basically little bags made of membrane that carry the wall material, so when they fuse together, their membranes form the plasma membranes of the new cells. The wall material gets dumped between the new membranes, forming the plant’s cell walls.
After cytokinesis is complete, the new cells move immediately into the G1 stage of interphase. No one stops to applaud the great accomplishment of successfully completing the mitosis process, which is really too bad because it’s the root of renewal and regeneration.
Figure 6-3:Cytokinesis.
Meiosis: It’s all about sex, baby
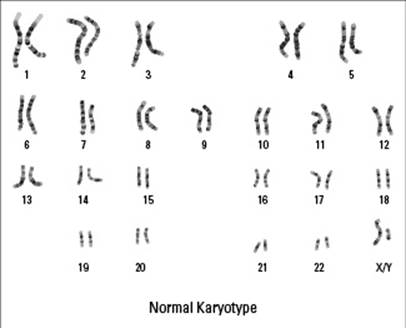
Meiosis is unique because the resulting cells have only half of their parent cells’ chromosomes, or singular pieces of DNA. Human body cells have 46 chromosomes, 2 each of 23 different kinds. The 23 pairs of chromosomes can be sorted by their physical similarities and lined up to form a chromosome map called a karyotype (see Figure 6-4). The two matched chromosomes in each pair are called homologous chromosomes (homo- means “same,” so these are chromosomes that have the same kind of genetic information). In each pair of your homologous chromosomes, one chromosome came from mom, and one came from dad. For every gene that your mom gave you, your dad also gave you a copy, so you have two copies of every gene (with the exception of genes on the X and Y chromosomes if you’re male).
The pairs of homologous chromosomes have the same kind of genetic information. If one of the two has a gene that affects eye color, for example, the other chromosome has the same gene in the same location. The messages in each gene may be slightly different — for example, one gene could have a message for light eyes whereas the other gene could have a message for dark eyes — but both chromosomes have the same type of gene in each location.
Figure 6-4: A human karyotype.
Human gametes (sperm cells and egg cells) have just 23 chromosomes. Through sexual reproduction (see Figure 6-5), a sperm and an egg join together to create a new individual, returning the chromosome number to 46. If gametes didn’t have half the genetic information, then the cell they form together, called a zygote, would have twice the normal genetic information for a human. And when gametes are produced, they can’t just get any 23 chromosomes — they have to get one of each pair of chromosomes. Otherwise, the zygote would have extras of some chromosomes and be missing others entirely. The resulting person wouldn’t have the correct genetic information and probably wouldn’t survive.
Figure 6-5: The human life cycle.
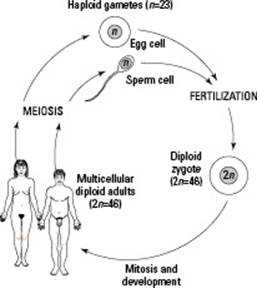
Meiosis is the type of cell division that separates chromosomes so gametes receive one of each type of chromosome. In humans, meiosis separates the 23 pairs of chromosomes so that each cell receives just one of each pair. Consequently, gametes have what’s known as a haploidnumber of chromosomes, or a single set. When the two gametes unite, they combine their chromosomes to reach the full complement of 46 chromosomes in a normal diploid cell (one with a double set of chromosomes, or two of each type).
Two cell divisions occur in meiosis, and the two halves of meiosis are called meiosis I and meiosis II.
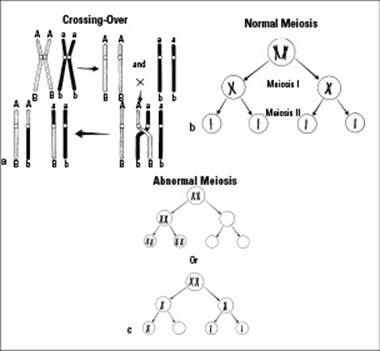
During meiosis I, homologous chromosomes are paired up and then separated into two daughter cells. Each daughter cell receives one of each chromosome pair, but the chromosomes are still replicated. (Remember that meiosis follows interphase, so DNA replication produced a copy of each chromosome. These two copies, called sister chromatids, are held together, forming replicated chromosomes. You can see that the chromosomes still look like X’s after meiosis I in Figure 6-6b.)
During meiosis II, the replicated chromosomes send one sister chromatid from each replicated chromosome to new daughter cells. After meiosis II, the four daughter cells each have one of each chromosome pair, and the chromosomes are no longer replicated. (Notice how the four daughter cells in Figure 6-6b don’t have sister chromatids?)
Figure 6-6:Crossing-over, meiosis, and nondisjunction.
In human males, meiosis takes place after puberty, when the diploid cells in the testes undergo meiosis to become haploid. In females, the process begins a lot earlier — in the fetal stage. While a little girl is dog-paddling around her mother’s womb, diploid cells complete the first part of meiosis and then migrate to the ovaries, where they hang out and wait until puberty. With the onset of puberty, the cells take turns entering meiosis II. (Just one per month; no pushing or shoving, please!) Usually one single egg cell is produced per cycle, although exceptions occur, which, if fertilized, lead to fraternal twins, or triplets, or quadruplets . . . you get the idea. The other meiotic cells simply disintegrate.
When a human sperm cell and a human egg cell — each with 23 chromosomes — unite in the process of fertilization, the diploid condition of the cell is restored. Further divisions by mitosis result in a complete human being.
The phases of meiosis are very similar to the phases of mitosis; they even have the same names, which can make distinguishing between the two rather tough. Just remember that the key difference between the phases of mitosis and meiosis is what’s happening to the number of chromosomes.
The next sections delve into the details of each phase of meiosis I and meiosis II.
Meiosis I
Meiosis I is the first step in sexual reproduction. The phases are as follows:
Prophase I: During this phase, the cell’s nuclear membrane breaks down, the chromatids coil to form visible chromosomes, the nucleoli break down and disappear, and the spindles form and attach to the chromosomes. But that’s not all. Prophase I is when something that’s absolutely critical to the successful separation of homologous chromosomes occurs: synapsis.
Synapsis happens when the two chromosomes of each pair find each other and stick together. The synapsis process begins when the homologous chromosomes move and lie next to each other. At this point, the two homologous chromosomes can swap equal amounts of DNA in an event called crossing-over (see Figure 6-6a). This swapping of materials results in four completely unique chromatids. This new arrangement of four chromatids is called a tetrad.
Crossing-over between homologous chromosomes during prophase I increases the genetic variability among gametes produced by the same organism. Every time meiosis occurs, crossing-over can happen a little differently, shuffling the genetic deck as gametes are made. This is one of the reasons that siblings can be so different from each other.
Metaphase I: This is when the pairs of homologous chromosomes line up in the center of the cell. The difference between metaphase I of meiosis and metaphase of mitosis is that the homologous pairs line up in the former, whereas individual chromosomes line up in the latter.
Anaphase I: During this phase, the two members of each homologous pair go to opposite sides of the cell, and the chromosome number is officially reduced from diploid to haploid.
Telophase I: This is when the cell takes a step back (or forward, depending on your perspective) to an interphase-like condition by reversing the events of prophase I. Specifically, the nuclear membrane reforms, the chromosomes uncoil and spread throughout the nucleus, the nucleoli reform, and the spindles break down.
Meiosis II
During meiosis II, both daughter cells produced by meiosis I continue their dance of division so that — in most cases — four gametes are the end result. The phases of meiosis II look very similar to the phases of meiosis I with one big exception: The cells start out with half the number of chromosomes as the original parent cell.
Meiosis II separates the sister chromatids of each replicated chromosome and sends them to opposite sides of the cell. Cells going from meiosis I to meiosis II don’t go through interphase again (after all, that would undo all the hard work of meiosis I).
Prophase II: As in mitosis’s prophase and meiosis’s prophase I, the nuclear membrane disintegrates, the nucleoli disappear, and the spindles form and attach to the chromosomes.
Metaphase II: Nothing too exciting here, folks. Just as in any old metaphase, the chromosomes line up at the equatorial plane. But remember that the number of chromosomes that lines up is half the number of the original parent cell (and half the number you’d see in mitosis).
Anaphase II: The sister chromatids of each replicated chromosome move away from each other to opposite sides of the cell.
Telophase II: The nuclear membrane and nucleoli reappear, the chromosomes stretch out for the briefest of rests, and the spindles disappear.
After meiosis II, it’s time for cytokinesis, which creates four haploid cells (which is impressive considering you had just one diploid cell at the beginning of meiosis).
How Sexual Reproduction Creates Genetic Variation
Sexual reproduction increases genetic variation in offspring, which in turn increases the genetic variability in species. You can see the effects of this genetic variability if you look at the children in a large family and note how each person is unique. Imagine this kind of variability expanded to include all the families you know (not to mention all the families of all the sexually reproducing organisms on Earth), and you begin to get a feel for the dramatic genetic impact of sexual reproduction.
The sections that follow familiarize you with some of the specific causes of genetic variation courtesy of meiosis and sexual reproduction.
Mutations
DNA polymerase occasionally makes uncorrected mistakes when copying a cell’s genetic information during DNA replication (which we walk you through earlier in this chapter). These mistakes are called spontaneous mutations, and they introduce changes into the genetic code. In addition, exposure of cells to mutagens (environmental agents, such as X-rays and certain chemicals, that cause changes in DNA) can increase the number of mutations that occur in cells. When changes occur in a cell that produces gametes, future generations are affected. (For more on the effect of mutations, see Chapter 8.)
Crossing-over
When homologous chromosomes come together during prophase I of meiosis, they exchange bits of DNA with each other. This crossing-over (illustrated in Figure 6-6a) results in new gene combinations and new chances for variety. Crossing-over is one way of explaining how a person can have red hair from his mother’s father and a prominent chin from his mother’s mother. After crossing-over, these two genes from two different people wound up together on the same chromosome in the person’s mother and got handed down together.
Independent assortment
Independent assortment occurs when homologous chromosomes separate during anaphase I of meiosis. When the homologous pairs of chromosomes line up in metaphase I, each pair lines up independently from the other pairs. So, the way the pairs are oriented during meiosis in one cell is different from the way they’re oriented in another cell. When the homologous chromosomes separate, many different combinations of homologous chromosomes can travel together toward the same side of the cell. How many different combinations of homologous chromosomes are possible in a human cell undergoing meiosis? Oh, just 223 — that’s 8,388,608 to be exact. Now you can begin to see why even large families can have many unique children.
Fertilization
Fertilization presents yet another opportunity for genetic diversity. Imagine millions of genetically different sperm swimming toward an egg. Fertilization is random, so the sperm that wins the race in one fertilization event is going to be different than the sperm that wins the next race. And, of course, each egg is genetically different too. So, fertilization produces random combinations of genetically diverse sperm and eggs, creating virtually unlimited possibilities for variation. That’s why every human being who has ever been born — and ever will be born — is genetically unique. Well, almost. Genetically identical twins can develop from the same fertilized egg, but even they can have subtle differences due to development.
Nondisjunction
Nothing’s perfect, even in the cellular world, which is why sometimes meiosis doesn’t occur quite right. When chromosomes don’t separate the way they’re supposed to, that’s called nondisjunction. The point of meiosis is to reduce the number of chromosomes from diploid to haploid, something that normally happens when homologous chromosomes separate from each other during anaphase I. Occasionally, however, a pair of chromosomes finds it just too hard to separate, and both members of the pair end up in the same gamete (see Figure 6-6c).
What happens next isn’t pretty. Two of the final four cells resulting from the meiotic process are missing a chromosome as well as the genes that chromosome carries. This condition usually means the cells are doomed to die. Each of the other two cells has an additional chromosome, along with the genetic material it carries. Well, that should be great for these cells, shouldn’t it? It should mean they’ll have an increased chance for genetic variation, and that’s a good thing, right?
Wrong! An extra chromosome is like an extra letter from the IRS. It’s not something to hope for. Many times these overendowed cells simply die, and that’s the end of the story. But sometimes they survive and go on to become sperm or egg cells. The real tragedy, then, is when an abnormal cell goes on to unite with a normal cell. When that happens, the resulting zygote (and offspring) has three of one kind of chromosome rather than the normal two. The term scientists use for this occurrence is trisomy.
Here’s the real problem with this scenario: All the cells that develop by mitosis to create the new individual will be trisomic (meaning they’ll have that extra chromosome). One possible abnormality occurring from an extra chromosome is Down syndrome, a condition that often results in some mental and developmental impairment and premature aging.
 Scientists have now pinpointed the chromosome related to Down syndrome; it’s Chromosome 21. If an egg with two number 21 chromosomes is fertilized with a normal sperm cell with just one number 21, the resulting offspring has 47 chromosomes (24 + 23 = 47), and Down syndrome occurs.
Scientists have now pinpointed the chromosome related to Down syndrome; it’s Chromosome 21. If an egg with two number 21 chromosomes is fertilized with a normal sperm cell with just one number 21, the resulting offspring has 47 chromosomes (24 + 23 = 47), and Down syndrome occurs.
You probably already know that the mother’s age is a factor in the creation of genetic abnormalities such as Down syndrome, but do you know why? The answer lies in the fact that meiosis I takes place in the fetal stage for females and then the cells rest in the ovaries until puberty, when one per month enters meiosis II in preparation for fertilization. If a cell has been waiting its turn for 40 or 45 years, it’s pretty darned old — in cellular terms at least. (Aging gametes aren’t such an issue for males because their sex cells don’t actually enter meiosis until after puberty. Meiosis is a continuous process for them, producing new cells all the time.)
Pink and blue chromosomes
Ever wish you could’ve been born the opposite sex so you wouldn’t have to spend so much of your budget on makeup or shave your face every morning? Sorry, but that was never really your decision to make. Like all other genetic characteristics, gender is determined at a chromosomal level.
In many organisms — including humans and fruit flies — the gender of an individual is determined by specific sex chromosomes, which scientists refer to as the X and Y chromosomes. The 23 pairs of human chromosomes can be divided into 22 pairs of autosomes, chromosomes that aren’t involved in the determination of gender, and one pair of sex chromosomes. Men and women have the same types of genes on the 22 autosomes and on the X chromosome. But only guys get a special gene, located on the Y chromosome, that jump-starts the formation of testes in boy fetuses when they’re about 6 weeks old. After the testes form, they produce testosterone, and it’s usually all boy from then on out. The Y chromosome is smaller than all the other chromosomes, but it packs one powerful little gene!
Monocultures, a threat to genetic diversity
Up until about the 1960s, most American farms were smaller operations that planted diverse crops and relied fairly heavily upon human labor to get the crops in. As farms started shifting to more mechanized processes, they grew larger and looked for greater standardization in crop plants. After all, machines work best at specialized, repetitive tasks. Farms started planting huge stands of monocultures, crop plants that are all genetically identical. This practice worries many scientists because they recognize the power and value of genetic diversity.
If an insect pest or fungal pathogen can successfully attack a crop plant, it has the potential to wipe out an entire monoculture at once and drastically impact the human food supply. In addition, large industrial farms increasingly rely upon artificial fertilizers and pesticides to support their specialized monocultures, which can upset the balance between the farm and the natural ecology.
As people start to realize that monoculture farming may have more dangers than benefits, some people are bucking the trend. Small farms are trying to make a comeback by producing diverse, organic crops, and companies are starting to collect older varieties of crop plants, calledheirloom varieties, and reintroduce them into the human food supply.