THE LIVING WORLD
Unit Eight. The Living Environment
37. Behavior and the Environment
37.11. Communication Within Social Groups
Many insects, fish, birds, and mammals live in social groups in which information is communicated between group members. For example, some individuals in mammalian societies serve as “guards.” When a predator appears, the guards give an alarm call, and group members respond by seeking shelter. Social insects, such as ants and honeybees, secrete chemicals called alarm pheromones that trigger attack behavior. Ants also deposit trail pheromones between the nest and a food source to lead other colony members to food (figure 37.13). Honeybees have an extremely complex dance language that directs nestmates to rich nectar sources.

Figure 37.13. Ants following a pheromone trail.
Trail pheromones organize cooperative foraging. The trails taken by the first ants to travel to a food source are soon followed by most of the other ants due to the release of pheromones by the first ants.
The Dance Language of the Honeybee
The European honeybee, Apis mellifera, lives in hives consisting of 30,000 to 40,000 individuals whose behaviors are integrated into a complex colony. Worker bees may forage miles from the hive, collecting nectar and pollen from a variety of plants on the basis of how energetically rewarding their food is. The food sources used by bees tend to occur in patches, and each patch offers much more food than a single bee can transport to the hive. A colony is able to exploit the resources of a patch because of the behavior of scout bees, which locate patches and communicate their location to hivemates through a dance language. Over many years, Nobel laureate Karl von Frisch was able to unravel the details of this communication system.
After a successful scout bee returns to the hive, she performs a remarkable behavior pattern called a waggle dance on a vertical comb (figure 37.14). The path of the bee during the dance resembles a figure-eight. On the straight part of the path, the bee vibrates or waggles her abdomen while producing bursts of sound. She may stop periodically to give her hivemates a sample of the nectar she has carried back to the hive in her crop. As she dances, she is followed closely by other bees, which soon appear as foragers at the new food source.
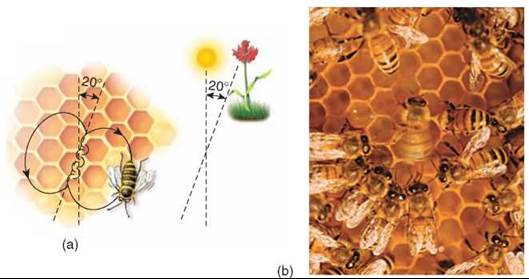
Figure 37.14. The waggle dance of honeybees.
(a) The angle between the food source, the nest, and the sun is represented by a dancing bee as the angle between the straight part of the dance and vertical. Here the food is 20 degrees to the right of the sun, and the straight part of the bee's dance on the hive is 20 degrees to the right of vertical. (b) A scout bee dancing in the hive.
Von Frisch and his colleagues claimed that the other bees use information in the waggle dance to locate the food source. According to their explanation, the scout bee indicates the direction of the food source by representing the angle between the food source, the hive, and the sun as the deviation from vertical of the straight part of the dance performed on the hive wall (that is, if the bee moved straight up, then the food source would be in the direction of the sun, but if the food was at a 30-degree angle relative to the sun’s position, then the bee would move upward at a 30-degree angle from vertical). The distance to the food source is indicated by the tempo, or degree of vigor, of the dance.
Adrian Wenner, a scientist at the University of California, did not believe that the dance language communicated anything about the location of food, and he challenged von Frisch’s explanation. Wenner maintained that flower odor was the most important cue allowing recruited bees to arrive at a new food source. A heated controversy ensued as the two groups of researchers published articles supporting their positions.
Such controversies can be very beneficial, because they often generate innovative experiments. In this case, the “dance language controversy” was resolved (in the minds of most scientists) in the mid-1970s by the creative research of James L. Gould. Gould devised an experiment in which hive members were tricked into misinterpreting the directions given by the scout bee’s dance. As a result, Gould was able to manipulate where the hive members would go if they were using visual signals. If odor was the cue they were using, hive members would have appeared at the food source, but instead they appeared exactly where Gould predicted. This confirmed von Frisch’s ideas.
Recently, researchers have extended the study of the honeybee dance language by building robot bees whose dances can be completely controlled. Their dances are programmed by a computer and perfectly reproduce the natural honeybee dance; the robots even stop to give food samples! The use of robot bees has allowed scientists to determine precisely which cues direct hivemates to food sources.
Primate Language
Some primates have a “vocabulary” that allows individuals to communicate the identity of specific predators. The vocalizations of African vervet monkeys, for example, distinguish eagles, leopards, and snakes. The two sonograms in figure 37.15b show the alarm calls for an eagle and a leopard, each eliciting different responses in other members of the troop. Chimpanzees and gorillas can learn to recognize a large number of symbols and use them to communicate abstract concepts.
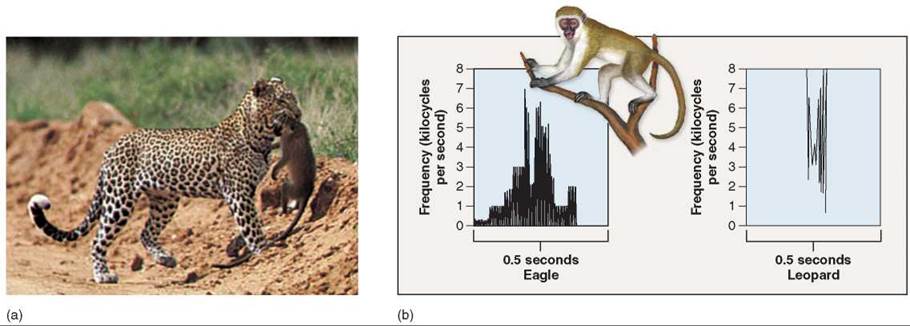
Figure 37.15. Primate semantics.
(a) This leopard, an efficient predator of primates, has attacked and will eat a vervet monkey. (b) Escaping a leopard attack presents a very different challenge to a vervet monkey than avoiding an eagle, another key predator of monkeys. Vervet monkeys give different alarm calls when leopards are sighted by troop members than they do when a member sees an eagle. Each distinctive call elicits a different and adaptive escape behavior.
The complexity of human language would at first appear to defy biological explanation, but closer examination suggests that the differences are in fact superficial—all languages share many basic structural similarities. All of the roughly 3,000 human languages draw from the same set of 40 consonant sounds (English uses two dozen of them), and any human can learn them. Researchers believe these similarities reflect the way our brains handle abstract information, a genetically determined characteristic of all humans.
Language develops at an early age in humans. Human infants are capable of recognizing the 40 consonant sounds characteristic of speech, including those not present in the particular language they will learn, while they ignore other sounds. In contrast, individuals who have not heard certain consonant sounds as infants can only rarely distinguish or produce them as adults. That is why English speakers have difficulty mastering the throaty French “r,” French speakers typically replace the English “th” with “z,” and native Japanese often substitute “r” for the unfamiliar English “l.” Children go through a “babbling” phase, in which they learn by trial and error how to make the sounds of language. Even deaf children go through a babbling phase using sign language. Next, children quickly and easily learn a vocabulary of thousands of words. Like babbling, this phase of rapid learning seems to be genetically programmed. It is followed by a stage in which children form simple sentences that, though they may be grammatically incorrect, can convey information. Learning the rules of grammar constitutes the final step in language acquisition.
While language is the primary channel of human communication, odor and other nonverbal signals (such as “body language”) may also convey information. However, it is difficult to determine the relative importance of these other communication channels in humans.
Key Learning Outcome 37.11. The study of animal communication involves analysis of the specificity of signals, their information content, and the methods used to produce and receive them.